Physiological and Proteomic Analyses Reveal Adaptive Mechanisms of Ryegrass (Annual vs. Perennial) Seedlings to Salt Stress


Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Stress Treatments
2.2. Fresh Weight and Dry Weight of the Seedlings
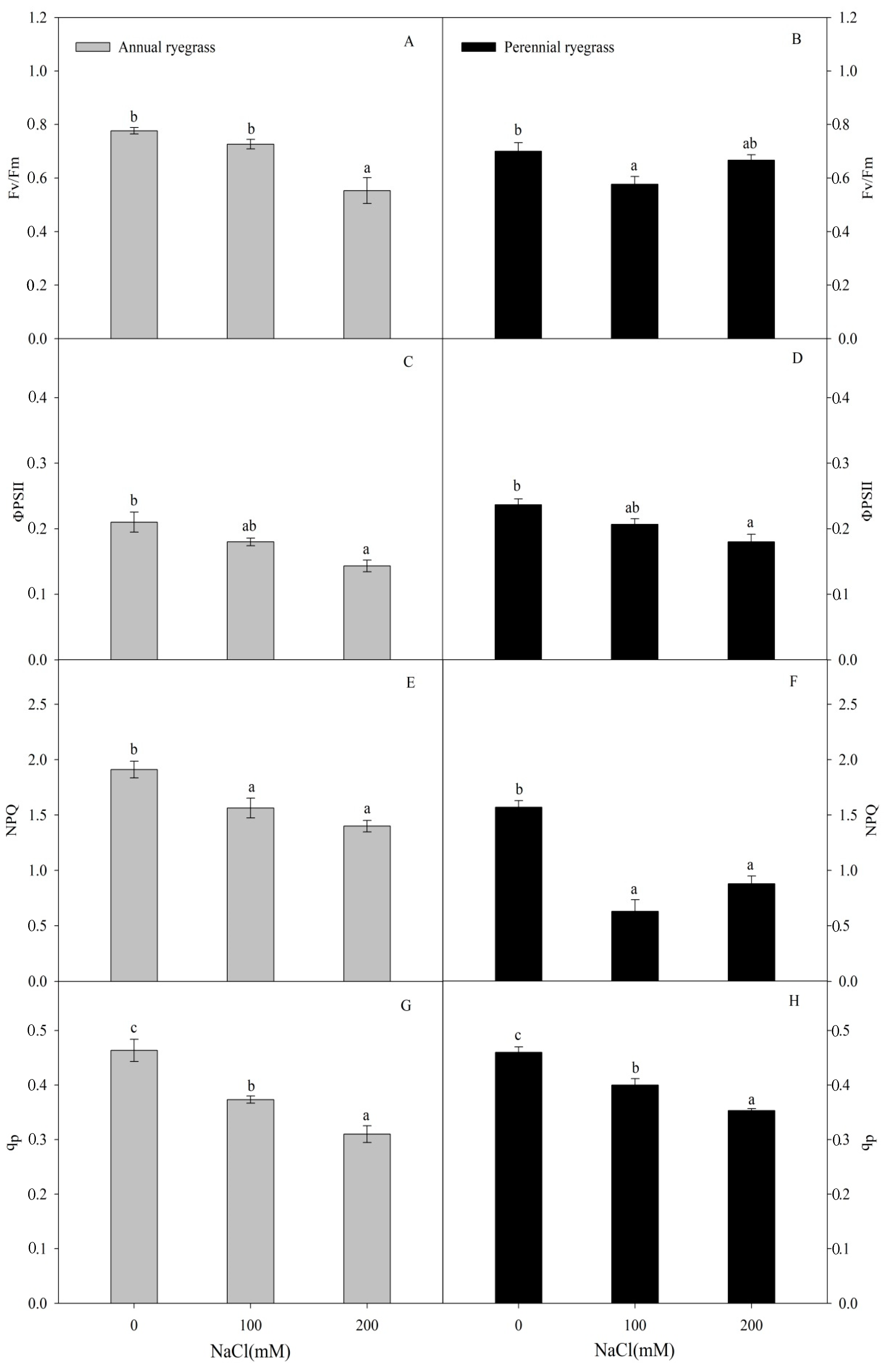
2.3. Chlorophyll Fluorescence
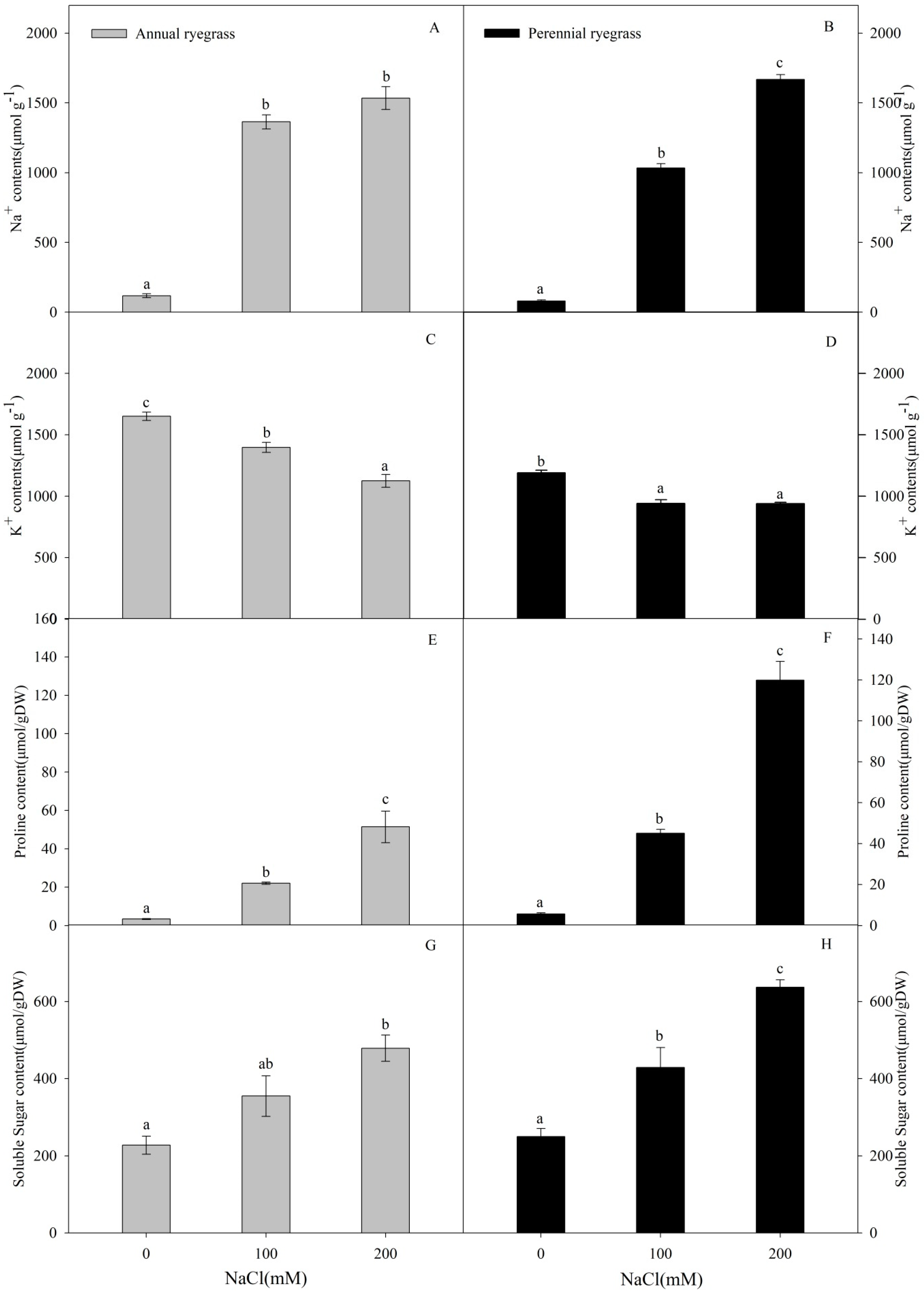
2.4. Inorganic Ion Contents
2.5. Proline and Total Soluble Sugar Contents
2.6. Protein Extraction
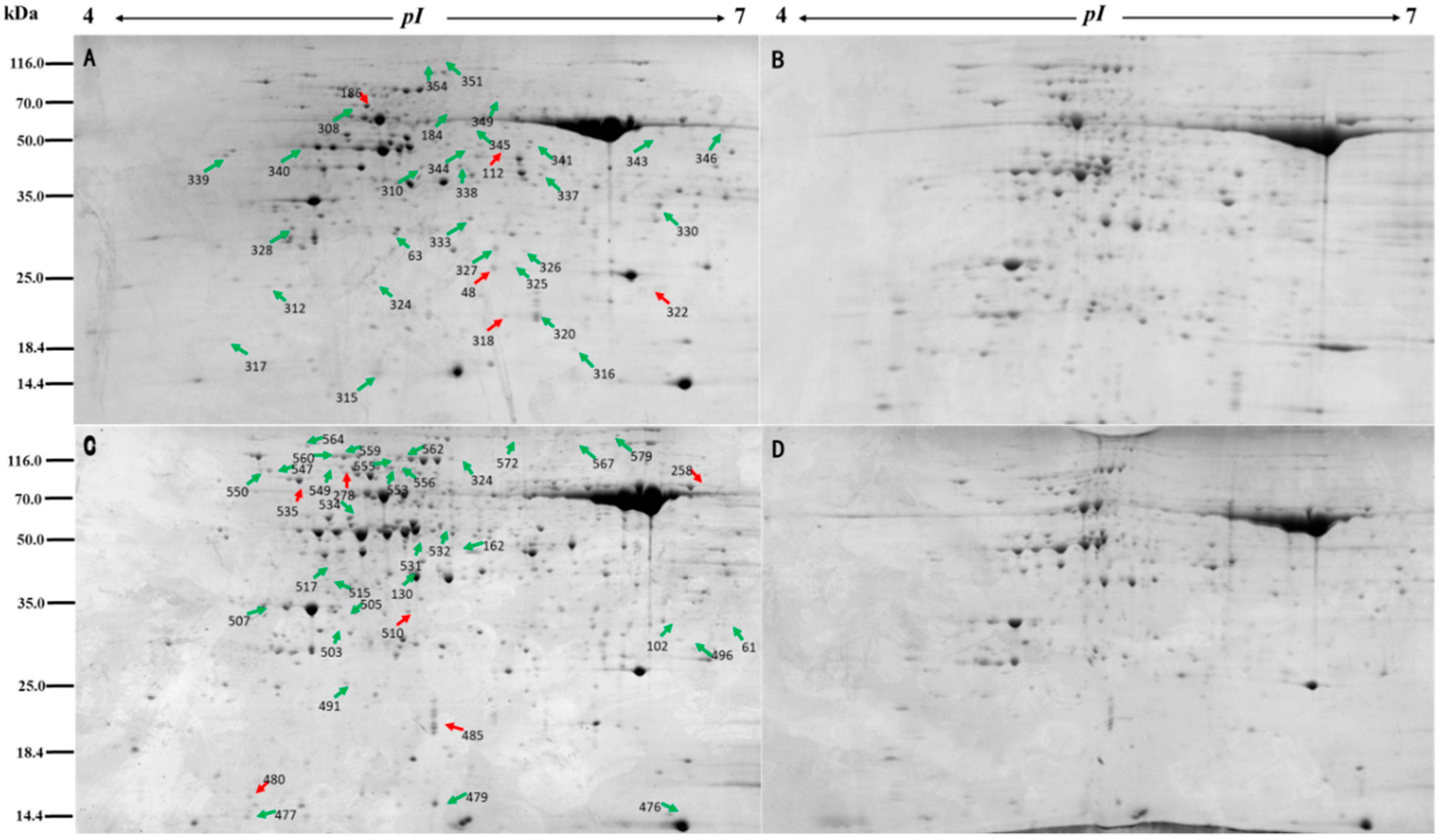
2.7. Two-Dimensional Electrophoresis
2.8. Gel Image Scanning and Protein Expression Abundance Analysis
2.9. In-Gel Digestion and MALDI-TOF/TOF Analysis
2.10. Statistical Analysis
3. Results
3.1. Physiological Changes of Annual Ryegrass and Perennial Ryegrass under Salinity
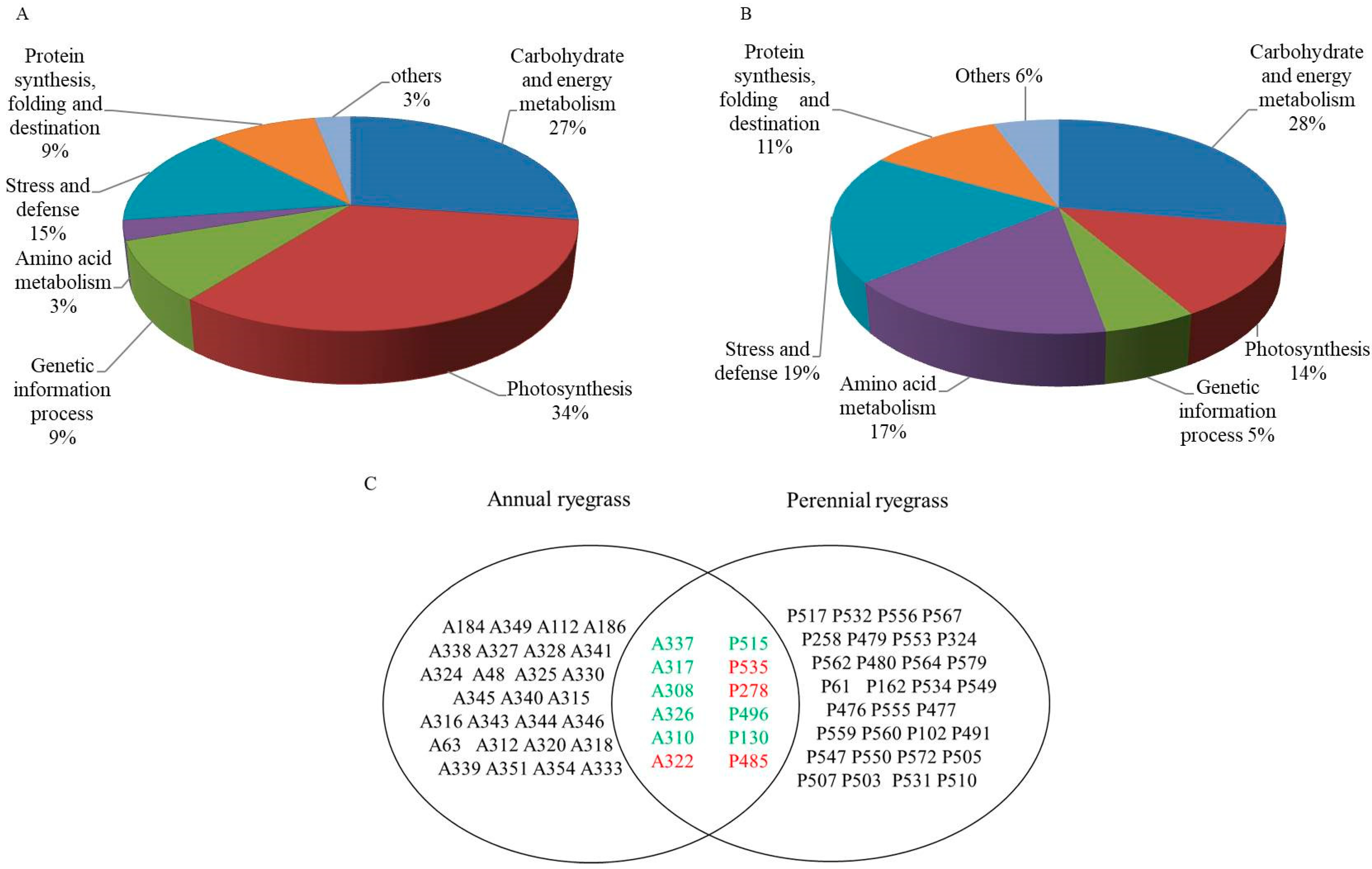
3.2. Proteomic Analysis of Annual Ryegrass and Perennial Ryegrass under Salinity
4. Discussion
4.1. Physiological Response of Annual and Perennial Ryegrass under Salinity Stress
4.2. Proteomics Analysis of Annual and Perennial Ryegrass under Salinity Stress
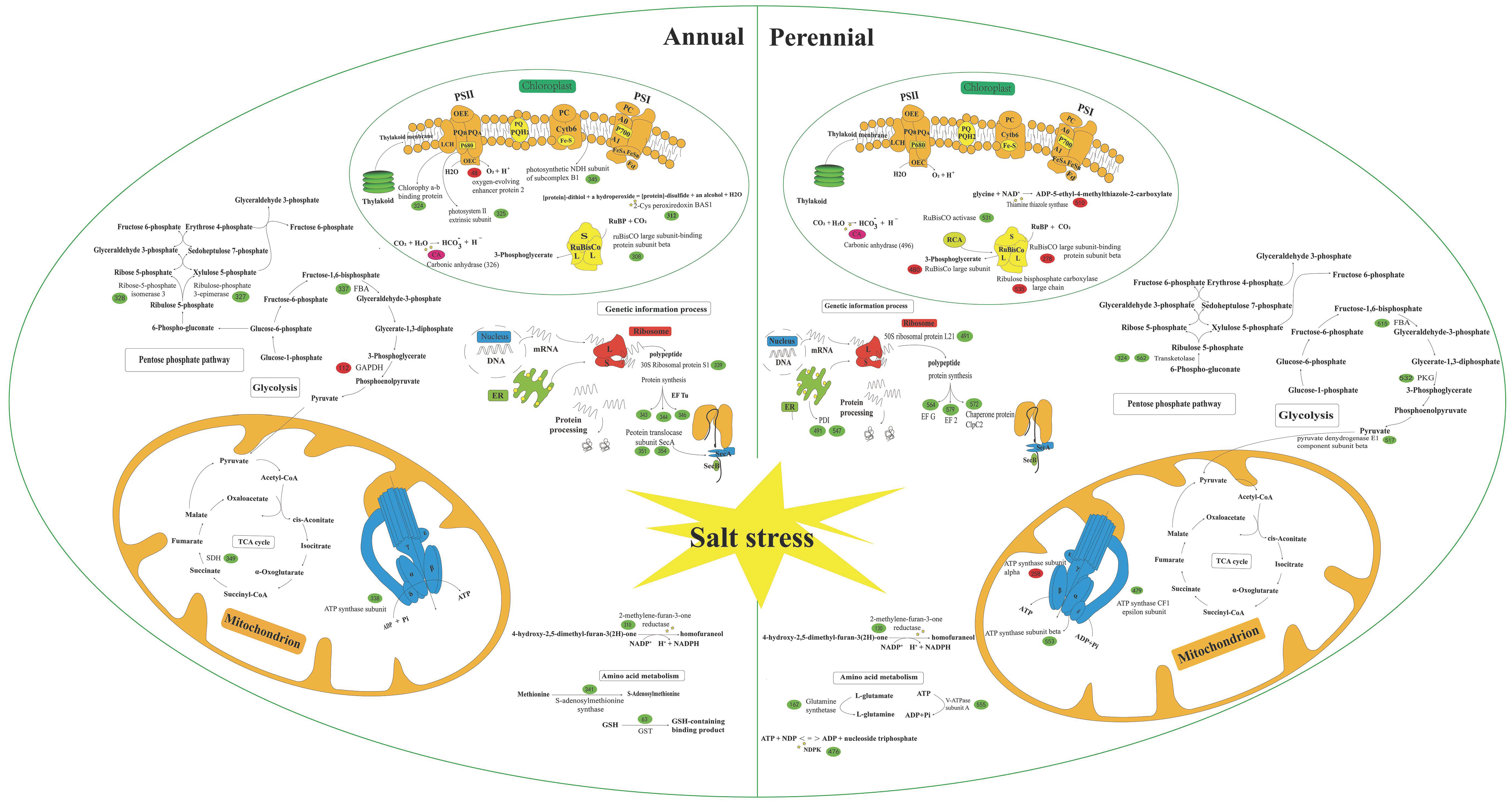
4.2.1. Carbohydrate and Energy Metabolism
4.2.2. Photosynthesis
4.2.3. Genetic Information Processes and Protein-Destiny
4.2.4. Amino Acid Metabolism and Stress Defense
5. Conclusions
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Humphreys, M.; Feuerstein, U.; Vandewalle, M.; Baert, J. Fodder Crops and Amenity Grasses; Boller, B., Posselt, F.U., Eds.; Veronesi: Dordrecht, The Netherlands, 2010; pp. 211–260. [Google Scholar]
- Pan, L.; Yang, Z.F.; Wang, J.P.; Wang, P.X.; Ma, X.; Zhou, M.L.; Li, J.; Nie, G.; Feng, G.Y.; Zhao, J.M.; et al. Comparative proteomic analyses reveal the proteome response to short-term drought in Italian ryegrass (Lolium multiflorum). PLoS ONE 2017, 12, e0184289. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.X.; Hua, X.Y.; Peng, X.Y.; Dong, B.L.; Yan, X.F. Germination Responses of Ryegrass (Annual vs. Perennial) Seed to the Interactive Effects of Temperature and Salt-Alkali Stress. Front. Plant Sci. 2018, 9, 1458. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Xu, Y.H.; Fei, S.; Li, L.J. Isolation, characterization and evolutionary analysis of resistance gene analogs in annual ryegrass, perennial ryegrass and their hybrid. Physiol. Plant. 2006, 126, 627–638. [Google Scholar] [CrossRef]
- Xiong, Y.W.; Fei, S.Z.; Arira, R.; Brummer, E.C.; Barker, R.E.; Jung, G.; Warnke, S.E. Identification of quantitative trait loci controlling winter hardiness in an annual×perennial ryegrass interspecific hybrid population. Mol. Breed. 2007, 19, 125–136. [Google Scholar] [CrossRef]
- Saddhe, A.A.; Malvankar, M.R.; Karle, S.B.; Kumar, K. Reactive nitrogen species: Paradigms of cellular signaling and regulation of salt stress in plants. Environ. Exp. Bot. 2019, 161, 86–97. [Google Scholar] [CrossRef]
- Agarwal, P.; Agarwal, P.K.; Gohil, D. Salinity Responses and Tolerance in Plants; Kumar, V., Wani, S., Suprasanna, P., Tran, L.S., Eds.; Springer: Cham, Switzerland, 2018; pp. 185–211. [Google Scholar]
- Fan, P.; Feng, J.; Jiang, P.; Chen, X.; Bao, H.; Nie, L.; Jiang, D.; Lv, S.; Kuang, T.; Li, Y. Coordination of carbon fixation and nitrogen metabolism in Salicornia europaea under salinity: Comparative proteomic analysis on chloroplast proteins. Proteomics 2011, 11, 4346–4367. [Google Scholar] [CrossRef] [PubMed]
- Dubouzet, J.G.; Sakuma, Y.; Ito, Y.; Kazuko, M.; Dubouzet, E.G.; Miura, S.; Seki, M.; Shinozaki, K.; Yamaguchi-Shinozaki, K. OsDREB genes in rice, Oryza sativa L., encode transcription activators that function in drought-, high-salt-and cold-responsive gene expression. Plant J. 2003, 33, 751–763. [Google Scholar] [CrossRef]
- Parida, A.K.; Das, A.B. Salt tolerance and salinity effects on plants: A review. Ecotox. Environ. Saf. 2005, 60, 324–349. [Google Scholar] [CrossRef]
- Li, M.N.; Zhang, K.; Long, R.C.; Sun, Y.; Kang, J.M.; Zhang, T.J.; Cao, S.H. iTRAQ-based comparative proteomic analysis reveals tissue-specific and novel early-stage molecular mechanisms of salt stress response in Carex rigescens. Environ. Exp. Bot. 2017, 143, 99–114. [Google Scholar] [CrossRef]
- Hu, T.; Li, H.Y.; Zhang, X.Z.; Luo, H.J.; Fu, J.M. Toxic effect of nacl on ion metabolism, antioxidative enzymes and gene expression of perennial ryegrass. Ecotox. Environ. Saf. 2011, 74, 2050–2056. [Google Scholar] [CrossRef]
- Hu, L.; Li, H.; Pang, H.; Fu, J. Responses of antioxidant gene, protein and enzymes to salinity stress in two genotypes of perennial ryegrass (Lolium perenne) differing in salt tolerance. J. Plant Physiol. 2012, 169, 0156. [Google Scholar] [CrossRef] [PubMed]
- Cao, L.; Wei, S.Q.; Han, L.; Qian, Y.Q.; Zhang, H.L.; Xin, H.B.; Sun, Z.Y. Gene cloning and expression of the pyrroline-5-carboxylate reductase gene of perennial ryegrass (Lolium perenne). Horticult. Plant J. 2015, 1, 113–120. [Google Scholar]
- Chen, Y.Y.; Lu, P.Z.; Sun, P.; Wei, L.; Chen, G.L.; Wu, D. Interactive salt—alkali stress and exogenous Ca2+, effects on growth and osmotic adjustment of, Lolium multiflorum, in a coastal estuary. Flora 2017, 229, 92–99. [Google Scholar] [CrossRef]
- Deshmukh, R.; Sonah, H.; Patil, G.; Chen, W.; Prince, S.; Mutava, R.; Vuong, T.; Valliyodan, B.; Nguyen, H.T. Integrating omic approaches for abiotic stress tolerance in soybean. Front. Plant Sci. 2014, 5, 244. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.N.; Peng, X.Y.; Salvato, F.; Wang, Y.C.; Yan, X.F.; Zhou, Z.Q.; Lin, J.X. Salt-adaptive strategies in oil seed crop Ricinus communis early seedlings (cotyledon vs. true leaf) revealed from proteomics analysis. Ecotox. Environ. Saf. 2019, 171, 12–25. [Google Scholar] [CrossRef]
- Yu, J.J.; Chen, S.X.; Zhao, Q.; Wang, T.; Yang, C.P.; Diaz, C.; Sun, G.R.; Dai, S.J. Physiological and proteomic analysis of salinity tolerance in Puccinellia Tenuiflora. J. Proteome Res. 2011, 10, 3852–3870. [Google Scholar] [CrossRef]
- Swami, A.K.; Alam, S.I.; Sengupta, N.; Sarin, R. Differential proteomic analysis of salt stress response in Sorghum bicolor leaves. Environ. Exp. Bot. 2011, 71, 321–328. [Google Scholar] [CrossRef]
- Cui, D.; Wu, D.; Liu, J.; Li, D.T.; Xu, C.Y.; Li, S.; Li, P.; Zhang, H.; Liu, X.; Jiang, C.; et al. Proteomic analysis of seedling roots of two maize inbred lines that differ significantly in the salt stress response. PLoS ONE 2015, 10, e0116697. [Google Scholar] [CrossRef]
- Pang, Q.Y.; Zhang, A.Q.; Zang, W.; Lei, W.; Yan, X.F. Integrated proteomics and metabolomics for dissecting the mechanism of global responses to salt and alkali stress in suaeda corniculata. Plant Soil. 2016, 402, 379–394. [Google Scholar] [CrossRef]
- Li, J.; Cui, G.; Hu, G.; Wang, M.; Zhang, P.; Qin, L. Proteome dynamics and physiological responses to short-term salt stress in Leymus chinensis leaves. PLoS ONE 2017, 12, e0183615. [Google Scholar] [CrossRef]
- Singh, R.P.; Runthala, A.; Khan, S.; Jha, P.N. Quantitative proteomics analysis reveals the tolerance of wheat to salt stress in response to enterobacter cloacae sbp-8. PLoS ONE 2017, 12, e0183513. [Google Scholar] [CrossRef] [Green Version]
- Jiang, Q.; Li, X.; Niu, F.; Sun, X.; Hu, Z.; Zhang, H. Itraq-based quantitative proteomic analysis of wheat roots in response to salt stress. Proteomics 2017, 17, 1600265. [Google Scholar] [CrossRef] [PubMed]
- Luo, M.; Zhao, Y.; Wang, Y.; Shi, Z.; Zhang, P.; Zhang, Y. Comparative proteomics of contrasting maize genotypes provides insights into salt-stress tolerance mechanisms. J. Proteome Res. 2017, 17, 141–153. [Google Scholar] [CrossRef]
- Dong, K.; Zhen, S.M.; Cheng, Z.W.; Cao, H.; Ge, P.; Yan, Y.M. Proteomic analysis reveals key proteins and phosphoproteins upon seed germination of wheat (Triticum aestivum L.). Front. Plant Sci. 2015, 6, 1017. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Li, Q.; Qi, Y.P.; Huang, W.L.; Yang, L.T.; Lai, N.W.; Ye, X.; Chen, L.S. Low pH-responsive proteins revealed by a 2-DE based MS approach and related physiological responses in Citrus leaves. BMC Plant Biol. 2008, 18, 188. [Google Scholar] [CrossRef] [PubMed]
- Jia, X.M.; Wang, H.; Svetla, S.; Zhu, Y.F.; Hu, Y.; Cheng, L.; Zhao, T.; Wang, Y.X. Comparative physiological responses and adaptive strategies of apple Malus halliana to salt, alkali and saline-alkali stress. Sci. Hortic. 2019, 245, 154–162. [Google Scholar] [CrossRef]
- Sun, L.L.; Xu, H.L.; Hao, H.D.; An, S.H.; Lu, C.T.; Wu, R.H.; Su, W.C. Effects of bensulfuron-methyl residue on photosynthesis and chlorophyll fluorescence in leaves of cucumber seedlings. PLoS ONE 2019, 14, e0215486. [Google Scholar] [CrossRef] [Green Version]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid determination of free proline for water stress studies. Plant Soil. 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Dubois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.A.; Smith, F. Colorimetric method for determination of sugars and related substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Yin, X.; He, D.; Gupta, R.; Yang, P. Physiological and proteomic analyses on artificially aged brassica napus seed. Front. Plant Sci. 2015, 6, 112. [Google Scholar]
- Sheffield, J.; Taylor, N.; Fauquet, C.; Chen, S. The cassava (Manihot esculenta Crantz) root proteome: Protein identification and differential expression. Proteomics 2006, 6, 1588–1598. [Google Scholar] [CrossRef]
- Yang, C.W.; Chong, J.N.; Li, C.Y.; Kim, C.M.; Shi, D.C.; Wang, D.L. Osmotic adjustment and ion balance traits of an alkali resistant halophyte Kochia sieversiana, during adaptation to salt and alkali conditions. Plant Soil. 2007, 294, 263–276. [Google Scholar] [CrossRef]
- Chaves, M.M.; Flexas, J.; Pinheiro, C. Photosynthesis under drought and salt stress: Regulation mechanisms from whole plant to cell. Ann. Bot. 2008, 103, 551–560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, Y.P.; Ding, X.T.; Zhang, D.; Deng, Q.; Yu, C.L.; Zhou, S.P.; Hui, D.F. Soil salinity increases the tolerance of excessive sulfur fumigation stress in tomato plants. Environ. Exp. Bot. 2017, 133, 70–77. [Google Scholar] [CrossRef] [Green Version]
- Mian, A.; Oomen, R.J.F.J.; Isayenkov, S.; Sentenac, H.; Maathuis, F.J.; Maathuis, F.J.M.; Véry, A.A. Over-expression of an Na+- and K+- permeable HKT transporter in barley improves salt tolerance. Plant J. 2011, 68, 468–479. [Google Scholar] [CrossRef]
- Li, X.Y.; Liu, J.J.; Zhang, Y.T.; Lin, J.X.; Mu, C.S. Physiological responses and adaptive strategies of wheat seedlings to salt and alkali stresses. Soil Sci. Plant Nutr. 2009, 55, 680–684. [Google Scholar] [CrossRef]
- Yang, C.W.; Xu, H.H.; Wang, L.L.; Liu, J.; Shi, D.C.; Wang, D.L. Comparative effects of salt-stress and alkali-stress on the growth, photosynthesis, solute accumulation, and ion balance of barley plants. Photosynthetica 2009, 47, 79–86. [Google Scholar] [CrossRef]
- Wang, X.; Fan, P.; Song, H.; Chen, X.; Li, X.; Li, Y. Comparative proteomic analysis of differentially expressed proteins in shoots of Salicornia europaea under different salinity. J. Proteome Res. 2009, 8, 3331–3345. [Google Scholar] [CrossRef]
- Swart, C.; Zhang, Y.; Kfm, B.; Bergmann, S.; Krahnert, I.; Nikoloski, Z. Protein-protein interactions and metabolite channeling in the plant tricarboxylic acid cycle. Nat. Commun. 2017, 8, 1–11. [Google Scholar]
- Chen, Z.; Sun, L.; Liu, P.; Liu, G.; Tian, J.; Liao, H. Malate synthesis and secretion mediated by a manganese-enhanced malate dehydrogenase confers superior manganese tolerance in Stylosanthes guianensis. Plant Physiol. 2015, 167, 176–188. [Google Scholar] [CrossRef] [Green Version]
- Kianipouya, A.; Roessner, U.; Jayasinghe, T.; Bazihizina, N. Epidermal bladder cells confer salinity stress tolerance in the halophyte quinoa and Atriplex species. Plant Cell Environ. 2017, 40, 1900–1915. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stincone, A.; Prigione, A.; Cramer, T.; Wamelink, M.M.C.; Campbell, K.; Cheung, E.; Olin-Sandoval, V.; Grüning, N.M.; Krüger, A.; Alam, M.T.; et al. The return of metabolism: Biochemistry and physiology of the pentose phosphate pathway. Biol. Rev. Camb. Philos. Soc. 2014, 90, 927–963. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Foyer, C.H.; Bloom, A.J.; Queval, G.; Noctor, G. Photorespiratory metabolism: Genes, mutants, energetics, and redox signaling. Ann. Rev. Plant Biol. 2009, 60, 455–484. [Google Scholar] [CrossRef] [PubMed]
- Kochetov, G.A.; Solovjeva, O.N. Sructure and functioning mechanism of transketolase. Ann. Rev. Plant Biol. 2014, 1844, 1608–1618. [Google Scholar]
- Hichem, H.; Naceur, A.E.; Mounir, D. Effects of salt stress on photosynthesis, PSII photochemistry and thermal energy dissipation in leaves oftwocorn (Zeamays, L.) varieties. Photosynthetica 2009, 47, 517–526. [Google Scholar] [CrossRef]
- Wang, J.Z.; Wang, J.; Wang, X.; Li, R.B.; Chen, B.S. Proteomic response of hybrid wild rice to cold stress at the seedling stage. PLoS ONE 2018, 13, e0198675. [Google Scholar] [CrossRef]
- Spreitzer, R.J.; Savucci, M.E. Rubisco: Structure, regulatory interactions and possibilities for a better enzyme. Ann. Rev. Plant Biol. 2002, 53, 449–485. [Google Scholar] [CrossRef] [Green Version]
- Ludwig, M. Evolution of carbonic anhydrase in C4 plants. Curr. Opin. Plant Biol. 2016, 31, 16–22. [Google Scholar] [CrossRef]
- Meng, X.J.; Zhao, Q.; Jin, Y.D.; Yu, J.J.; Yin, Z.P.; Chen, S.X.; Dai, S.J. Chilling-responsive mechanisms in halophyte puccinellia tenuiflora seedlings revealed from proteomics analysis. J. Proteom. 2016, 143, 365–381. [Google Scholar] [CrossRef]
- Meiri, E.; Levitan, A.; Guo, F.; Christopher, D.A.; Schaefer, D.; Zryd, J.-P.; Danon, A. Characterization of three PDI-like genes in Physcomitrella patens and construction of knock-out mutants. Mol. Genet. Genom. 2002, 267, 231–240. [Google Scholar]
- Askari, H.; Edqvist, J.; Hajheidari, M.; Kafi, M.; Salekdeh, G.H. Effects of salinity levels on proteome of Suaeda aegyptiaca leaves. Proteomics 2006, 6, 2542–2554. [Google Scholar] [CrossRef] [PubMed]
- Alam, I.; Sharmin, S.A.; Kim, K.H.; Yang, J.K.; Choi, M.S.; Lee, B.H. Proteome analysis of soybean roots subjected to short-term drought stress. Plant Soil. 2010, 333, 491–505. [Google Scholar] [CrossRef]
- Denison, F.C.; Paul, A.L.; Zupanska, A.K.; Ferl, R.J. 14-3-3 proteins in plant physiology. Semin. Cell Dev. Biol. 2011, 22, 720–727. [Google Scholar] [CrossRef]
- Yu, H.T.; Wang, T. Proteomic dissection of endosperm starch granule associated proteins reveals a network coordinating starch biosynthesis and amino acid metabolism and glycolysis in rice endosperms. Front. Plant Sci. 2016, 7, 707. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.B.; Zhao, J.; He, X.Y.; Sun, H.Y.; Zhang, G.P.; Wu, F.B. Comparative proteomic analysis of drought tolerance in the two contrasting Tibetan wild genotypes and cultivated genotype. BMC Genom. 2015, 16, 432. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruiz, J.; Blumwald, E. Salinity-induced glutathione synthesis inbrassica napus. Planta (Berlin) 2002, 214, 965–969. [Google Scholar] [CrossRef]
- Stotz, H.U.; Long, S.R. Expression of the pea (pisum sativum L.) α-tubulin gene tuba1 is correlated with cell division activity. Plant Mol. Biol. 1999, 41, 601–614. [Google Scholar] [CrossRef] [PubMed]
- Assmann, S.M. G proteins go green: A plant G protein signaling FAQ sheet. Science 2005, 310, 71–73. [Google Scholar] [CrossRef] [Green Version]
- Lv, D.W.; Zhu, G.R.; Zhu, D.; Bian, Y.W.; Liang, X.N.; Cheng, Z.W.; Deng, X.; Yan, Y.M. Proteomic and phosphoproteomic analysis reveals the response and defense mechanism in leaves of diploid wheat T. monoccum under salt stress and recovery. J. Proteom. 2016, 143, 93–105. [Google Scholar] [CrossRef]
- Carter, C.; Thornburg, R.W. Germin-like proteins: Structure, phylogeny and function. J. Plant Biol. 1999, 42, 97–108. [Google Scholar] [CrossRef]
- Tan-Wong, S.M.; Zaugg, J.B.; Camblong, J.; Xu, X.Y.; Zhang, D.W.; Mischo, H.E.; Ansari, A.Z.; Luscombe, N.M.; Steinmetz, L.M.; Proudfoot, N.J. Gene loops enhance transcriptional directionality. Science 2012, 338, 671–675. [Google Scholar] [CrossRef] [Green Version]
- Wyers, F.; Rougemaille, M.; Badis, G.; Rousselle, J.C.; Dufour, M.E.; Boulay, J.; Regnault, B.; Devaux, F.; Namane, A.; Seraphin, B.; et al. Cryptic pol II transcripts are degraded by a nuclear quality control pathway involving a new poly(A) polymerase. Cell 2005, 121, 725–737. [Google Scholar] [CrossRef] [Green Version]
- Peinado-Guevara, L.I.; López-Meyer, M.; López-Valenzuela, J.A.; Maldonado-Mendoza, I.E.; Galindo-Flores, H.; Campista-León, S.; Medina, S. Comparative proteomic analysis of leaf tissue from tomato plants colonized with rhizophagus irregularis. Symbiosis 2017, 73, 93–106. [Google Scholar] [CrossRef]
- Xu, T.D.; Wen, M.Z.; Nagawa, S.; Fu, Y.; Chen, J.G.; Wu, M.J.; Perrot-Rechenmann, C.; Friml, J.; Jones, A.M.; Yang, Z.B. Cell surface- and rho GTPase-based auxin signaling controls cellular interdigitation in Arabidopsis. Cell 2010, 143, 99–110. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Annual Ryegrass | Perennial Ryegrass | |||||
|---|---|---|---|---|---|---|
| Control | 100 mM | 200 mM | Control | 100 mM | 200 mM | |
| Fresh weight (g·plant−1) | 0.3295 ± 0.0196 c | 0.2243 ± 0.0134 b | 0.1432 ± 0.0139 a | 0.2484 ± 0.0098 b | 0.1223 ± 0.0122 a | 0.0905 ± 0.0113 a |
| Dry weight (g·plant−1) | 0.0412 ± 0.0012 c | 0.0296 ± 0.0036 b | 0.0221 ± 0.0020 a | 0.0301 ± 0.0014 c | 0.0198 ± 0.0016 b | 0.0211 ± 0.0034 a |
| Spot No. a | Accession No. b | Protein Name | Plant Species c | Thero kD/pI d | Score e | Sequence Coverage f | Peptides Matched g | Fold Changes | p-Value |
|---|---|---|---|---|---|---|---|---|---|
| Carbohydrate and Energy Metabolism (9) | |||||||||
| TCA Cycle | |||||||||
| 184 | I1IYJ8 | Betaine aldehyde dehydrogenase | Brachypodium distachyon | 53.35/5.46 | 63 | 7 | 1 | −2.1 | 0.05 |
| 349 | A0A287E122 | Succinate dehydrogenase | Hordeum vulgare subsp. vulgare | 57.21/5.96 | 264 | 11 | 3 | −2.08 | 0.03 |
| Glycolysis | |||||||||
| 112 | A0A0Q3LKL4 | Glyceraldehyde-3-phosphate dehydrogenase | Brachypodium distachyon | 47.65/6.03 | 371 | 12 | 3 | 3.8 | 0.02 |
| 337 | A0A3B6RIU4 | Fructose-bisphosphate aldolase, cytosolic | Triticum aestivum | 41.46/5.85 | 62 | 3 | 1 | −2.55 | 0.01 |
| 186 | K6ZY00 | Phosphoglycerate mutase | Phleum pratense | 32.25/5.68 | 299 | 14 | 2 | 1.68 | 0.04 |
| ATP Synthesis | |||||||||
| 338 | A0A2X0TKP9 | ATP synthase subunit | Triticum aestivum | 40.12/8.13 | 417 | 21 | 4 | −1.83 | 0.01 |
| Pentose Phosphate Pathway | |||||||||
| 327 | A0A287PP03 | Ribulose-phosphate 3-epimerase | Hordeum vulgare subsp. vulgare | 29.21/8.29 | 508 | 25 | 4 | −2.23 | 0.01 |
| 328 | A0A2X0TSQ1 | Ribose-5-phosphate isomerase 3 | Triticum aestivum | 29.62/6.34 | 600 | 30 | 4 | −1.62 | 0.05 |
| 341 | A0A077RWS5 | S-adenosylmethionine synthase | Triticum aestivum | 43.28/5.61 | 512 | 17 | 4 | −1.76 | 0.04 |
| Photosynthesis (11) | |||||||||
| Photosynstem Ⅱ | |||||||||
| 324 | A0A164ZG13 | Chlorophyll a-b binding protein | Daucus carota subsp. sativus | 26.49/5.96 | 61 | 4 | 1 | −3.57 | 0.01 |
| 325 | A0A0E0AJY3 | Photosystem II extrinsic subunit | Oryza glumipatula | 27.35/8.56 | 111 | 11 | 1 | −1.53 | 0.03 |
| 48 | A0A287I5I5 | Oxygen-evolving enhancer protein 2 | Hordeum vulgare subsp. vulgare | 20.68/5.75 | 231 | 21 | 2 | 1.67 | 0.01 |
| Electron Transfer Chain | |||||||||
| 345 | I1I2I7 | Photosynthetic NDH subunit of subcomplex B1 | Brachypodium distachyon | 52.12/6.36 | 148 | 8 | 1 | −1.92 | 0.05 |
| Calvin Cycle | |||||||||
| 340 | A0A067L8Y9 | Ribulose bisphosphate carboxylase/oxygenase activase | Jatropha curcas | 52.21/5.56 | 284 | 10 | 3 | −1.68 | 0.01 |
| 315 | Q9SP00 | Ribulose-1,5-bisphosphate carboxylase/oxygenase small chain | Avena agadiriana | 19.08/8.59 | 250 | 20 | 2 | −1.37 | 0.02 |
| 316 | Q9SP00 | Ribulose-1,5-bisphosphate carboxylase/oxygenase small chain | Avena agadiriana | 19.08/8.59 | 73 | 7 | 1 | −1.1 | 0.01 |
| 317 | U5YI33 | Ribulose-1,5-bisphosphate carboxylase/oxygenase large chain | Sarcochlamys pulcherrima | 25.79/6.33 | 204 | 8 | 1 | −1.63 | 0.03 |
| 330 | P0C510 | Ribulose-1.5-bisphosphate carboxylase/oxygenase large chain | Oryza sativa | 53.42/0.51 | 447 | 12 | 3 | −2.1 | 0.03 |
| 308 | A0A0D3GC12 | Ribulose-1.5-bisphosphate carboxylase/oxygenase large subunit-binding protein subunit beta | Oryza barthii | 64.3/5.6 | 201 | 7 | 2 | −4.94 | 0.01 |
| 326 | A0A0Q3GCH7 | Carbonic anhydrase | Brachypodium distachyon | 30.33/6.81 | 422 | 24 | 4 | −2.55 | 0.01 |
| Genetic Information Process (3) | |||||||||
| 343 | A0A0J7YLM1 | Elongation factor Tu (Fragment) | Beta vulgaris subsp. vulgaris | 44.45/5.24 | 117 | 4 | 1 | −1.63 | 0.01 |
| 344 | A0A0J7YLM1 | Elongation factor Tu (Fragment) | Beta vulgaris subsp. vulgaris | 44.45/5.24 | 92 | 4 | 1 | −1.39 | 0.04 |
| 346 | F8SVL8 | Elongation factor 1 (Fragment) | Phalaenopsis hybrid cultivar | 44.29/9.19 | 207 | 14 | 1 | −1.96 | 0.01 |
| Amino Acid Metabolism (1) | |||||||||
| 63 | A0A165FYU2 | Glutathione-S-transferase | Lolium perenne | 25.57/5.71 | 342 | 23 | 3 | −2.63 | 0.04 |
| Stress and Defense (5) | |||||||||
| 312 | I1IAA5 | 2-Cys peroxiredoxin BAS1 | Brachypodium distachyon | 28.51/6.43 | 190 | 41 | 4 | −1.85 | 0.02 |
| 310 | I1I6Q9 | 2-methylene-furan-3-one reductase | Brachypodium distachyon | 39.84/8.29 | 233 | 10 | 2 | −1.57 | 0.02 |
| 320 | A0A1E5VJG5 | Germin-like protein 8-14 | Dichanthelium oligosanthes | 21.89/5.69 | 249 | 16 | 2 | −1.77 | 0.04 |
| 318 | A0A059BYP6 | Auxin-binding protein ABP19a | Eucalyptus grandis | 21.76/8.93 | 183 | 11 | 1 | 2.04 | 0.01 |
| 322 | A0A059BYP6 | Auxin-binding protein ABP19a | Eucalyptus grandis | 21.76/8.93 | 119 | 11 | 1 | 1.9 | 0.01 |
| Protein Synthesis, Folding and Destination (3) | |||||||||
| 339 | A0A287P2Y9 | 30S ribosomal protein S1 | Hordeum vulgare subsp. vulgare | 443.45/4.67 | 264 | 13 | 4 | −3.64 | 0.01 |
| 351 | A0A2K2D841 | Protein translocase subunit SecA | Brachypodium distachyon | 111.44/5.82 | 476 | 9 | 6 | −2.59 | 0.01 |
| 354 | A0A0D3EN60 | Protein translocase subunit SecA | Oryza barthii | 111.53/5.79 | 55 | 3 | 1 | −1.73 | 0.02 |
| Others (1) | |||||||||
| 333 | A0A0D3EUB5 | Hypothetical protein OsI_03835 | Oryza barthii | 56.86/9.09 | 74 | 5 | 1 | −2.96 | 0.02 |
| Spot No a | Accession No b | Protein Name | Plant Species c | Thero kD/pI d | Score e | Sequence Coverage f | Petides Matched g | Fold Changes | p-Value |
|---|---|---|---|---|---|---|---|---|---|
| Carbohydrate and Energy Metabolism (10) | |||||||||
| Glycolysis | |||||||||
| 515 | A0A287QT87 | Fructose-bisphosphate aldolase, cytosolic | Hordeum vulgare subsp. vulgare | 41.85/6.39 | 131 | 8 | 2 | −1.24 | 0.01 |
| 517 | A0A0D3H2L3 | Pyruvate dehydrogenase E1 component subunit beta | Oryza barthii | 40.18/5.35 | 342 | 13 | 4 | −1.48 | 0.01 |
| 532 | I1HI25 | Phosphoglycerate kinase | Brachypodium distachyon | 50.19/6.99 | 598 | 17 | 4 | −1.77 | 0.02 |
| 556 | A0A397Y6S9 | Probable phosphoglucomutase | Brassica campestris | 63.61/5.84 | 147 | 3 | 1 | −1.53 | 0.01 |
| 567 | A0A0D3GDS1 | Sucrose synthase | Oryza barthii | 92.61/5.96 | 82 | 2 | 1 | −4.79 | 0.01 |
| ATP Synthesis | |||||||||
| 258 | A0A2L0VAS4 | ATP synthase subunit alpha | Lamarckia aurea | 55.76/6.11 | 815 | 22 | 7 | 1.64 | 0.05 |
| 479 | K4PA35 | ATP synthase CF1 epsilon subunit | Lolium multiflorum | 15.26/5.25 | 612 | 71 | 5 | −2.21 | 0.01 |
| 553 | A9L9Y6 | ATP synthase subunit beta | Festuca arundinacea | 52.99/5.17 | 211 | 15 | 1 | −2.21 | 0.01 |
| Pentose Phosphate Pathway | |||||||||
| 324 | A0A194YGV9 | Transketolase | Sorghum bicolor | 79.92/6.00 | 242 | 6 | 2 | −1.27 | 0.05 |
| 562 | A0A194YGV9 | Transketolase | Sorghum bicolor | 79.92/6.00 | 216 | 6 | 2 | −1.84 | 0.01 |
| Photosynthesis (5) | |||||||||
| 278 | A0A287VF44 | Ribulose-1.5-bisphosphate carboxylase/oxygenase large subunit-binding protein subunit beta | Hordeum vulgare subsp. vulgare | 59.69/5.02 | 587 | 16 | 6 | 2.43 | 0.02 |
| 480 | A0A0U1XKS7 | Ribulose-1,5-bisphosphate carboxylase/oxygenase large subunit | Polypogon fugax | 2.978/4.08 | 93 | 53 | 1 | 1.89 | 0.01 |
| 531 | A0A067L8Y9 | Ribulose-1,5-bisphosphate carboxylase/oxygenase activase | Jatropha curcas | 52.21/5.56 | 330 | 10 | 3 | −1.73 | 0.01 |
| 535 | A0A075M0L1 | Ribulose-1,5-bisphosphate carboxylase large chain | Brachiaria serrata | 50.08/6.6 | 570 | 20 | 5 | 1.34 | 0.01 |
| 496 | A0A059Q285 | Carbonic anhydrase | Saccharum hybrid cultivar | 22.77/7.6 | 66 | 8 | 1 | −2.56 | 0.01 |
| Genetic Information Process (2) | |||||||||
| 564 | A0A0Q3E7E5 | Elongation factor G | Brachypodium distachyon | 84.23/5.25 | 869 | 17 | 7 | −2.86 | 0.01 |
| 579 | A0A067JM98 | Elongation factor 2 | Jatropha curcas | 94.99/5.9 | 267 | 9 | 3 | −2.64 | 0.04 |
| Amino Acid Metabolism (6) | |||||||||
| 61 | A0A0D3ER48 | GTP-binding nuclear protein | Oryza barthii | 25.33/6.38 | 57 | 14 | 1 | −2.76 | 0.02 |
| 162 | C5IW60 | Glutamine synthetase | Lolium perenne | 38.97/5.59 | 164 | 9 | 1 | −1.37 | 0.04 |
| 534 | Q0WV25 | Tubulin alpha-4 chain | Arabidopsis thaliana | 50.19/4.93 | 91 | 8 | 1 | −1.53 | 0.01 |
| 549 | A0A287R7K7 | Serine/threonine-protein phosphatase 2A 65 kDa regulatory subunit A beta isoform | Hordeum vulgare subsp. vulgare | 66.23/4.95 | 417 | 18 | 3 | −1.92 | 0.01 |
| 476 | Q9LKM0 | Nucleoside diphosphate kinase | Lolium perenne | 16.49/6.3 | 441 | 33 | 4 | −2.56 | 0.01 |
| 555 | A0A341Y4U2 | vacuolar proton-ATPase subunit A | Triticum aestivum | 68.09/5.19 | 521 | 13 | 5 | −4.58 | 0.02 |
| Stress and Defense (7) | |||||||||
| 130 | A0A3B6NMG8 | 2-methylene-furan-3-one reductase | Triticum aestivum | 40.01/7.63 | 315 | 9 | 2 | −1.4 | 0.02 |
| 477 | A0A3B6KDF0 | Thioredoxin M-type | Triticum aestivum | 19.66/8.67 | 106 | 9 | 1 | −1.68 | 0.03 |
| 510 | A0A368Q639 | Thiamine thiazole synthase | Setaria italica | 40.87/6.00 | 272 | 12 | 2 | 3.48 | 0.04 |
| 559 | A0A3B6KDM7 | Probable mediator of RNA polymerase II transcription subunit 37e | Triticum aestivum | 71.43/5.07 | 930 | 20 | 8 | −2.86 | 0.01 |
| 560 | A0A3B6LHE6 | Probable mediator of RNA polymerase II transcription subunit 37e | Triticum aestivum | 71.43/5.07 | 556 | 13 | 5 | −1.67 | 0.02 |
| 102 | A0A0D9VL60 | Annexin | Leersia perrieri | 31.81/6.77 | 81 | 9 | 1 | −2.74 | 0.02 |
| 485 | A0A059BYP6 | auxin-binding protein ABP19a | Eucalyptus grandis | 21.76/8.93 | 164 | 11 | 1 | 1.75 | 0.01 |
| Protein Synthesis, Folding and Destination (4) | |||||||||
| 491 | A0A287U0T7 | 50S ribosomal protein L21 | Hordeum vulgare subsp. vulgare | 25.87/9.63 | 260 | 24 | 3 | −1.68 | 0.03 |
| 547 | A0A024FR39 | Protein disulfide-isomerase | Triticum aestivum | 53.99/5.1 | 200 | 9 | 1 | −2.55 | 0.02 |
| 550 | A0A0D3HJ84 | Protein disulfide-isomerase | Oryza barthii | 57.06/4.92 | 86 | 3 | 1 | −1.53 | 0.01 |
| 572 | A0A3B6KF51 | Chaperone protein ClpC2 | Triticum aestivum | 102.03/6.55 | 423 | 9 | 5 | −1.73 | 0.04 |
| Signal Transduction (1) | |||||||||
| 507 | A0A0D9ZCC0 | 14-3-3-like protein GF14-F | Oryza glumipatula | 29.24/4.81 | 448 | 28 | 4 | −4.63 | 0.01 |
| Others (2) | |||||||||
| 503 | A0A0D9X940 | Haloacid dehalogenase-like hydrolase domain-containing protein At4g39970 | Leersia perrieri | 35.38/6.23 | 216 | 11 | 2 | −1.95 | 0.03 |
| 505 | A0A1D5UQE8 | Haloacid dehalogenase-like hydrolase domain-containing protein At4g39970 | Triticum aestivum | 34.99/5.89 | 314 | 20 | 3 | −2.35 | 0.01 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Peng, X.; Yu, D.; Yan, J.; Zhang, N.; Lin, J.; Wang, J. Physiological and Proteomic Analyses Reveal Adaptive Mechanisms of Ryegrass (Annual vs. Perennial) Seedlings to Salt Stress. Agronomy 2019, 9, 843. https://doi.org/10.3390/agronomy9120843
Peng X, Yu D, Yan J, Zhang N, Lin J, Wang J. Physiological and Proteomic Analyses Reveal Adaptive Mechanisms of Ryegrass (Annual vs. Perennial) Seedlings to Salt Stress. Agronomy. 2019; 9(12):843. https://doi.org/10.3390/agronomy9120843
Chicago/Turabian StylePeng, Xiaoyuan, Dafu Yu, Junxin Yan, Na Zhang, Jixiang Lin, and Jinghong Wang. 2019. "Physiological and Proteomic Analyses Reveal Adaptive Mechanisms of Ryegrass (Annual vs. Perennial) Seedlings to Salt Stress" Agronomy 9, no. 12: 843. https://doi.org/10.3390/agronomy9120843