Effects of Alternative Administration Programs of a Synbiotic Supplement on Broiler Performance, Foot Pad Dermatitis, Caecal Microbiota, and Blood Metabolites

 , , , ,
, , , ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Management
2.2. Experimental Diets and Synbiotic Treatments
2.3. Productive Performance and Evaluation of FPD
2.4. Blood and Caecal Content Collections
2.5. Plasma Metabolomics Analyses
2.6. DNA Extraction Protocol
2.7. 16S rRNA Amplicon Sequencing
2.8. Statistical Analysis of Productive Performance and FPD
2.9. Statistical Analysis of Caecal Bacteria Relative Abundances and of Plasma Metabolomics
3. Results
3.1. Productive Performance and FPD
3.2. Plasma Metabolome
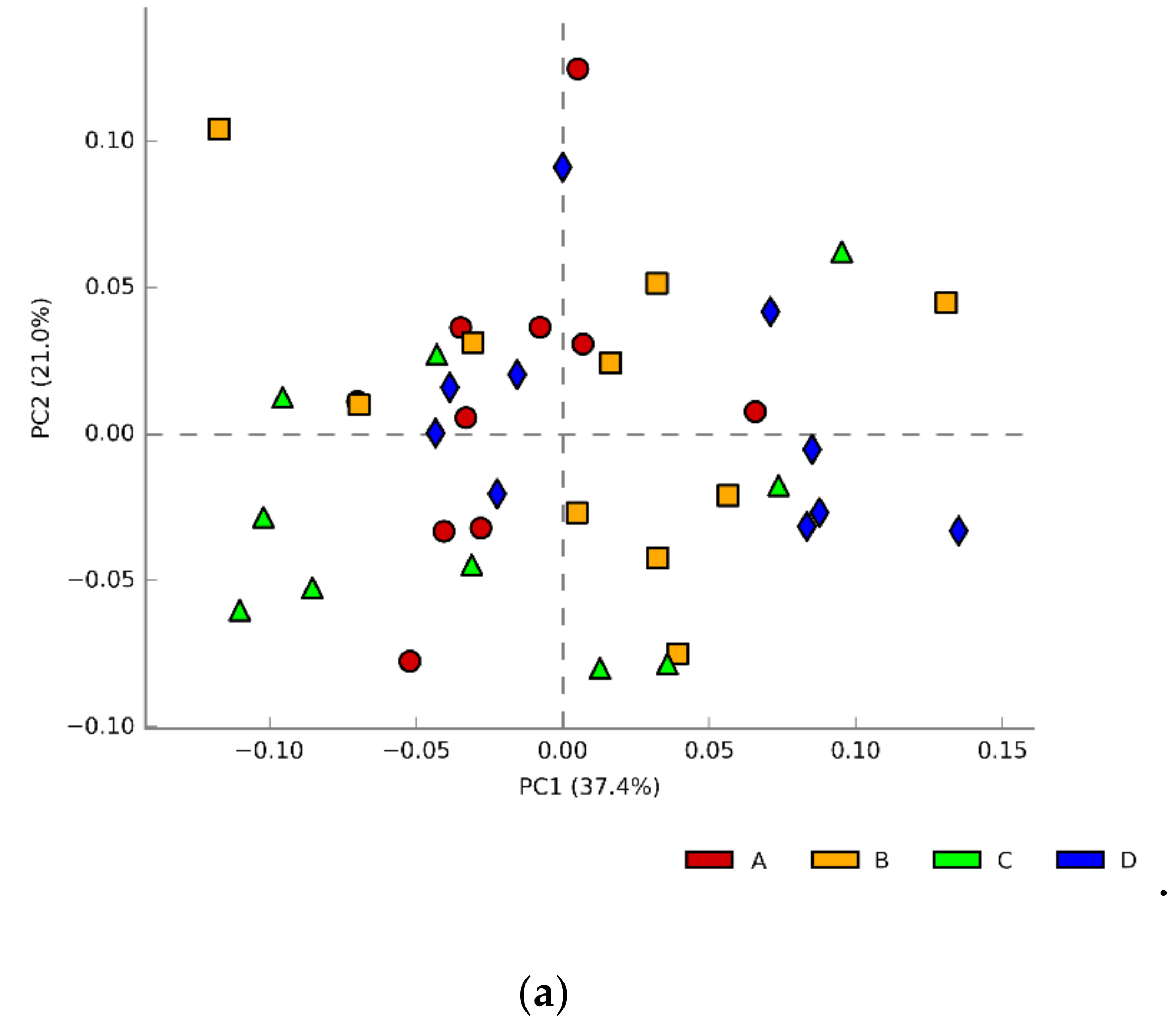
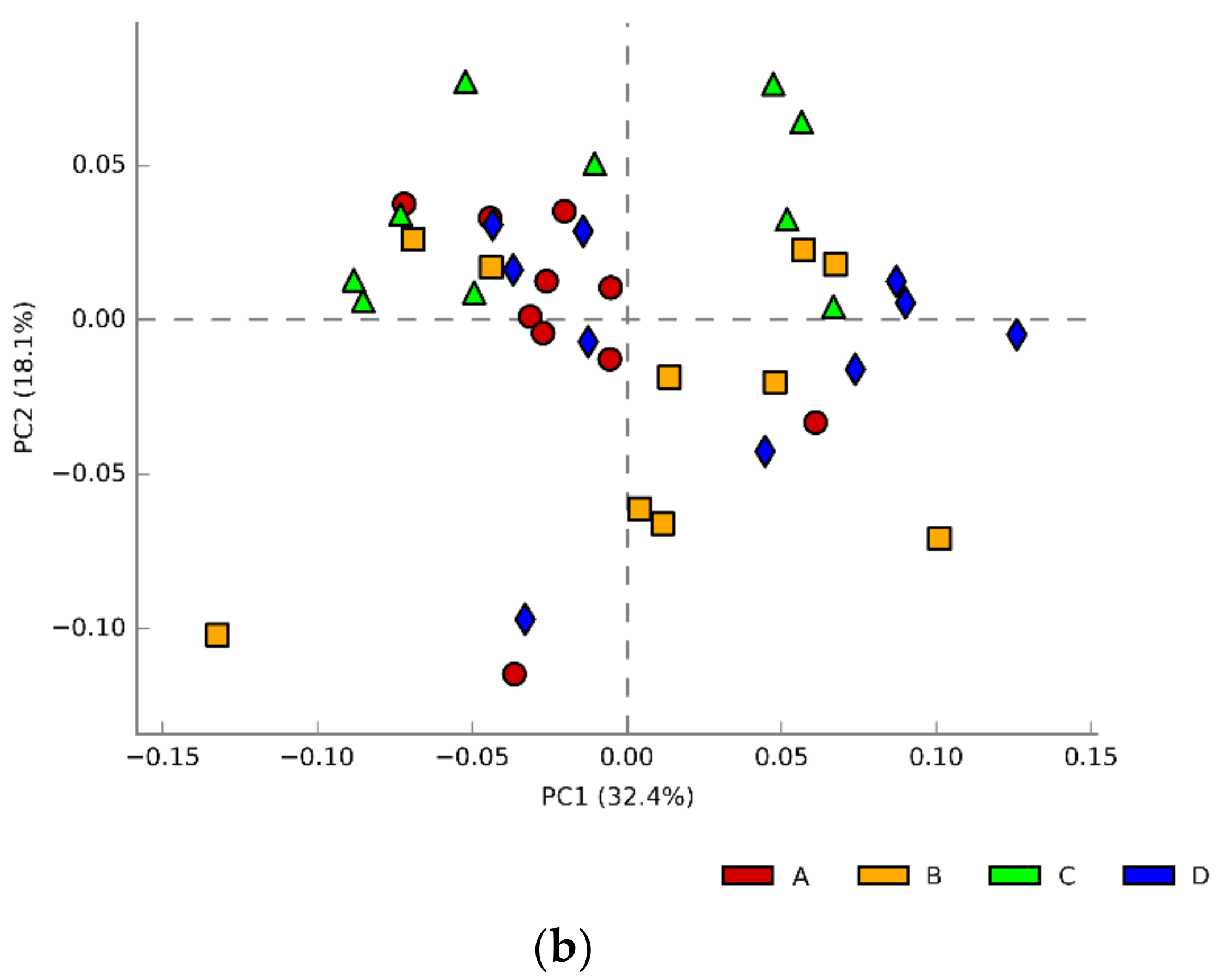
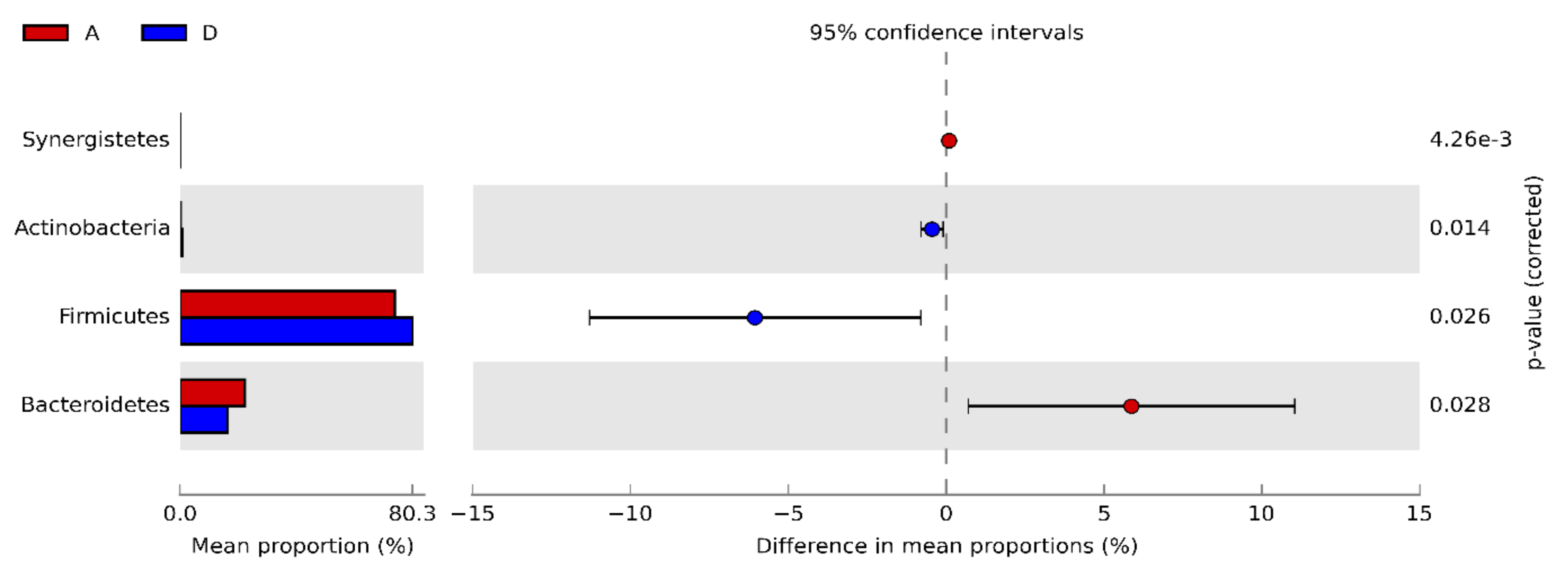
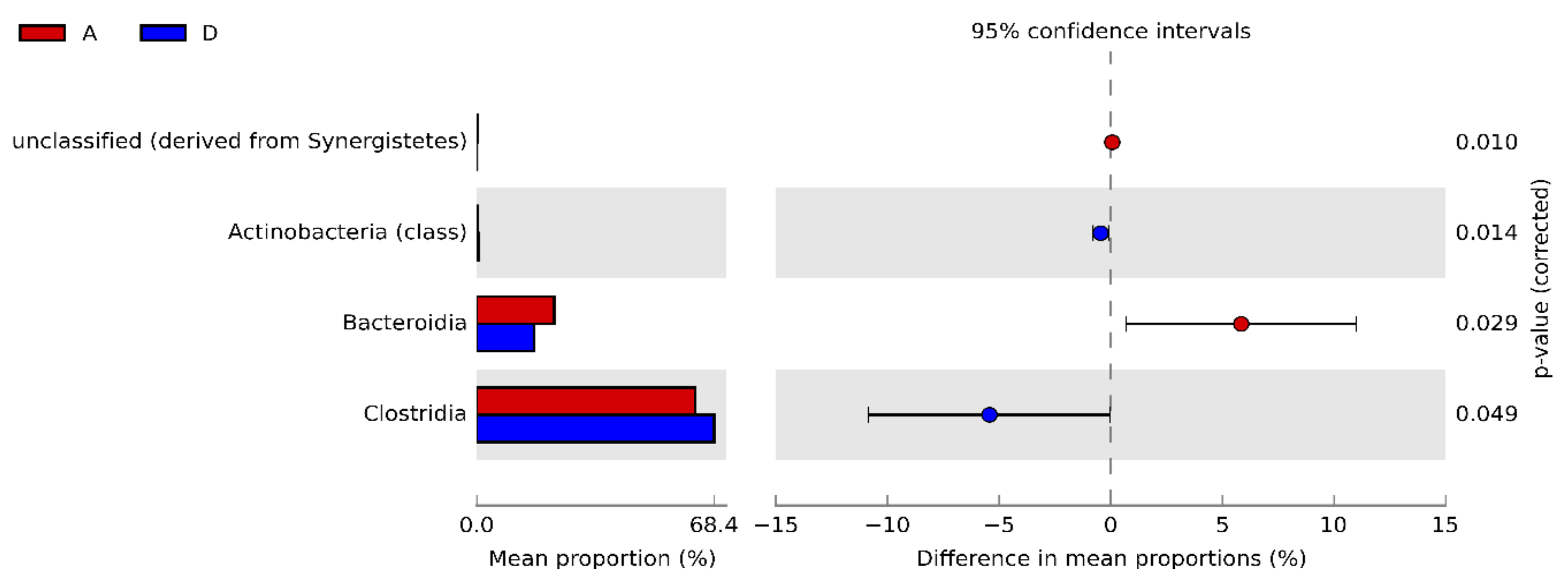
3.3. Caecal Microbiota
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Oviedo-Rondón, E.O. Holistic view of intestinal health in poultry. Anim. Feed Sci. Technol. 2019, 250, 1–8. [Google Scholar] [CrossRef]
- Kogut, M. Gut health in poultry. CAB Rev. Perspect. Agric. Vet. Sci. Nutr. Nat. Resour. 2017, 12. [Google Scholar] [CrossRef]
- Kogut, M.H.; Arsenault, R.J. Editorial: Gut Health: The new paradigm in food animal production. Front. Vet. Sci. 2016, 3, 10–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Celi, P.; Cowieson, A.J.; Fru-Nji, F.; Steinert, R.E.; Kluenter, A.-M.; Verlhac, V. Gastrointestinal functionality in animal nutrition and health: New opportunities for sustainable animal production. Anim. Feed Sci. Technol. 2017, 234, 88–100. [Google Scholar] [CrossRef]
- Moore, P.R.; Evenson, A.; Luckey, T.D.; Mccoy, E.; Elvehjem, C.A.; Hart, E.B. Use of sulfasuxidine, streptothricin, and streptomycin in nutritional studies with the chick. J. Biol. Chem. 1946, 165, 437–441. [Google Scholar]
- Jukes, T.H.; Stokstad, E.L.R.; Tayloe, R.R.; Cunha, T.J.; Edwards, H.M.; Meadows, G.B. Growth-promoting effect of aureomycin on pigs. Arch. Biochem. 1950, 26, 324–325. [Google Scholar]
- Woolhouse, M.; Ward, M.; van Bunnik, B.; Farrar, J. Antimicrobial resistance in humans, livestock and the wider environment. Philos. Trans. R. Soc. B 2015, 370, 20140083. [Google Scholar] [CrossRef]
- O’Neill, J.; Tackling Drug-Resistance Infections Globally: Final Report and Recommendations. The review on Antimicrobial Resistance. Available online: https://amr-review.org/sites/default/files/160518_Finalpaper_withcover.pdf (accessed on 26 November 2019).
- European Legislation. Regulation (EC) No 1831/2003 of the European Parliament and of the Council of 22 September 2003 on Additives for Use in Animal Nutrition. Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/PDF/?uri=CELEX:32003R1831&from=EN (accessed on 25 October 2019).
- Flynn, D. South Korea Bans Antibiotics in Animal Feed. Available online: https://www.foodsafetynews.com/2011/06/south-korea-bans-antibiotics-in-animal-feed/#.Uh99Nm2KLm0 (accessed on 27 November 2019).
- Xiao, Y. A National Action Plan to Contain Antimicrobial Resistance in China: Contents, Actions and Expectations. Available online: http://resistancecontrol.info/2017/a-national-action-plan-to-contain-antimicrobial-resistance-in-china-contents-actions-and-expectations/ (accessed on 27 November 2019).
- Millen, D.D.; Pacheco, R.D.L.; Meyer, P.M.; Rodrigues, P.H.M.; De Beni Arrigoni, M. Current outlook and future perspectives of beef production in Brazil. Anim. Front. 2011, 1, 46–52. [Google Scholar] [CrossRef] [Green Version]
- Millet, S.; Maertens, L. The European ban on antibiotic growth promoters in animal feed: From challenges to opportunities. Vet. J. 2011, 187, 143–144. [Google Scholar] [CrossRef]
- Gaggìa, F.; Mattarelli, P.; Biavati, B. Probiotics and prebiotics in animal feeding for safe food production. Int. J. Food Microbiol. 2010, 141, S15–S28. [Google Scholar] [CrossRef]
- Tayeri, V.; Seidavi, A.; Asadpour, L.; Phillips, C.J.C. A comparison of the effects of antibiotics, probiotics, synbiotics and prebiotics on the performance and carcass characteristics of broilers. Vet. Res. Commun. 2018, 42, 195–207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Y.; Iji, P.A.; Choct, M. Dietary modulation of gut microflora in broiler chickens: A review of the role of six kinds of alternatives to in-feed antibiotics. Worlds Poult. Sci. J. 2009, 65, 97–114. [Google Scholar] [CrossRef]
- Gadde, U.; Kim, W.H.; Oh, S.T.; Lillehoj, H.S. Alternatives to antibiotics for maximizing growth performance and feed efficiency in poultry: A review. Anim. Health Res. Rev. 2017, 18, 26–45. [Google Scholar] [CrossRef] [PubMed]
- Sugiharto, S. Role of nutraceuticals in gut health and growth performance of poultry. J. Saudi Soc. Agric. Sci. 2016, 15, 99–111. [Google Scholar] [CrossRef] [Green Version]
- Miles, R.D.; Bootwalla, S.M. Direct-fed microbials in animal production. In Direct-fed Microbials in Animal Production. A Review of Literature; Hutcheson, D.P., Ed.; National Feed Ingredient Association: West Des Moines, IA, USA, 1991; pp. 117–132. [Google Scholar]
- Cox, N.A.; Bailey, J.S.; Blankenship, L.C.; Gildersleeve, R.P. Research note: In ovo administration of a competitive exclusion culture treatment to broiler embryos. Poult. Sci. 1992, 71, 1781–1784. [Google Scholar] [CrossRef] [PubMed]
- Goren, E.; de Jong, W.A.; Doornenbal, P.; Koopman, J.P.; Kennis, H.M. Protection of chicks against salmonella infection induced by spray application of intestinal microflora in the hatchery. Vet. Q. 1984, 6, 73–79. [Google Scholar] [CrossRef]
- Madej, J.P.; Stefaniak, T.; Bednarczyk, M. Effect of in ovo-delivered prebiotics and synbiotics on lymphoid-organs’ morphology in chickens. Poult. Sci. 2015, 94, 1209–1219. [Google Scholar] [CrossRef]
- Ritzi, M.M.; Abdelrahman, W.; Van-Heerden, K.; Mohnl, M.; Barrett, N.W.; Dalloul, R.A. Combination of probiotics and coccidiosis vaccine enhances protection against an Eimeria challenge. Vet. Res. 2016, 47, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Pender, C.M.; Kim, S.; Potter, T.D.; Ritzi, M.M.; Young, M.; Dalloul, R.A. In ovo supplementation of probiotics and its effects on performance and immune-related gene expression in broiler chicks. Poult. Sci. 2017, 96, 1052–1062. [Google Scholar] [CrossRef]
- Baldwin, S.; Hughes, R.J.; Van, T.T.H.; Moore, R.J.; Stanley, D. At-hatch administration of probiotic to chickens can introduce beneficial changes in gut microbiota. PLoS ONE 2018, 13, e0194825. [Google Scholar] [CrossRef]
- European Legislation. Council Directive 2007/43/EC of 28 June 2007 Laying down Minimum Rules for the Protection of Chickens Kept for Meat Production. Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/?uri=celex%3A32007L0043 (accessed on 25 October 2019).
- European Legislation. Council Regulation (EC) No 1099/2009 of 24 September 2009 on the Protection of Animals at the Time of Killing. Available online: https://eur-lex.europa.Eu/legal-content/EN/ALL/?uri=celex%3A32009R1099 (accessed on 25 October 2019).
- European Legislation. Directive 2010/63/EU of the European Parliament and of the Council of 22 September 2010 on the Protection of Animals Used for Scientific Purposes. Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/?uri=celex%3A32010L0063 (accessed on 25 October 2019).
- Ekstrand, C.; Carpenter, T.E.; Andersson, I.; Algers, B. Prevalence and control of foot-pad dermatitis in broilers in Sweden. Br. Poult. Sci.. 1998, 39, 318–324. [Google Scholar] [CrossRef] [PubMed]
- Ventrella, D.; Laghi, L.; Barone, F.; Elmi, A.; Romagnoli, N.; Bacci, M.L. Age-related 1H NMR characterization of Cerebrospinal Fluid in newborn and young healthy piglets. PLoS ONE 2016, 11. [Google Scholar] [CrossRef] [PubMed]
- Barbara, G.; Scaioli, E.; Barbaro, M.R.; Biagi, E.; Laghi, L.; Cremon, C.; Marasco, G.; Colecchia, A.; Picone, G.; Salfi, N.; et al. Gut microbiota, metabolome and immune signatures in patients with uncomplicated diverticular disease. Gut 2017, 66, 1252–1261. [Google Scholar] [CrossRef] [PubMed]
- Kneen, M.A.; Annegarn, H.J. Algorithm for fitting XRF, SEM and PIXE X-ray spectra backgrounds. Nucl. Instrum. Methods Phys. Res. Sect. B Beam Interact. Mater. Atoms 1996, 109–110, 209–213. [Google Scholar] [CrossRef]
- Liland, K.H.; Almøy, T.; Mevik, B.H. Optimal choice of baseline correction for multivariate calibration of spectra. Appl. Spectrosc. 2010, 64, 1007–1016. [Google Scholar] [CrossRef]
- Dieterle, F.; Ross, A.; Schlotterbeck, G.; Senn, H. Probabilistic quotient normalization as robust method to account for dilution of complex biological mixtures. Application in1H NMR metabonomics. Anal. Chem. 2006, 78, 4281–4290. [Google Scholar] [CrossRef]
- Wishart, D.S.; Tzur, D.; Knox, C.; Eisner, R.; Guo, A.C.; Young, N.; Cheng, D.; Jewell, K.; Arndt, D.; Sawhney, S.; et al. HMDB: The human metabolome database. Nucleic Acids Res. 2007, 35. [Google Scholar] [CrossRef]
- De Cesare, A.; Sirri, F.; Manfreda, G.; Moniaci, P.; Giardini, A.; Zampiga, M.; Meluzzi, A. Effect of dietary supplementation with Lactobacillus acidophilus D2/CSL (CECT 4529) on caecum microbioma and productive performance in broiler chickens. PLoS ONE 2017, 12. [Google Scholar] [CrossRef] [Green Version]
- Meyer, F.; Paarmann, D.; D’Souza, M.; Olson, R.; Glass, E.; Kubal, M.; Paczian, T.; Rodriguez, A.; Stevens, R.; Wilke, A.; et al. The metagenomics RAST server—A public resource for the automatic phylogenetic and functional analysis of metagenomes. BMC Bioinform. 2008, 9, 386. [Google Scholar] [CrossRef] [Green Version]
- Pruesse, E.; Quast, C.; Knittel, K.; Fuchs, B.M.; Ludwig, W.; Peplies, J.; Glöckner, F.O. SILVA: A comprehensive online resource for quality checked and aligned ribosomal RNA sequence data compatible with ARB. Nucleic Acids Res. 2007, 35, 7188–7196. [Google Scholar] [CrossRef] [Green Version]
- DeSantis, T.Z.; Hugenholtz, P.; Larsen, N.; Rojas, M.; Brodie, E.L.; Keller, K.; Huber, T.; Dalevi, D.; Hu, P.; Andersen, G.L. Greengenes, a chimera-checked 16S rRNA gene database and workbench compatible with ARB. Appl. Environ. Microbiol. 2006, 72, 5069–5072. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cole, J.R.; Wang, Q.; Cardenas, E.; Fish, J.; Chai, B.; Farris, R.J.; Kulam-Syed-Mohideen, A.S.; McGarrell, D.M.; Marsh, T.; Garrity, G.M.; et al. The Ribosomal Database Project: Improved alignments and new tools for rRNA analysis. Nucleic Acids Res. 2009, 37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parks, D.H.; Tyson, G.W.; Hugenholtz, P.; Beiko, R.G. STAMP: Statistical analysis of taxonomic and functional profiles. Bioinformatics 2014, 30, 3123–3124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blankenship, L.C.; Bailey, J.S.; Cox, N.A.; Stern, N.J.; Brewer, R.; Williams, O. Two-step mucosal competitive exclusion flora treatment to diminish salmonellae in commercial broiler chickens. Poult. Sci. 1993, 72, 1667–1672. [Google Scholar] [CrossRef]
- Chen, M.; Stern, N.J.; Bailey, J.S.; Cox, N.A. Administering mucosal competitive exclusion flora for control of salmonellae. J. Appl. Poult. Res. 1998, 7, 384–391. [Google Scholar] [CrossRef]
- Roberfroid, M.B. Prebiotics and synbiotics: Concepts and nutritional properties. Br. J. Nutr. 1998, 80, S197–S202. [Google Scholar] [CrossRef] [Green Version]
- Torok, V.A.; Hughes, R.J.; Mikkelsen, L.L.; Perez-Maldonado, R.; Balding, K.; MacAlpine, R.; Percy, N.J.; Ophel-Keller, K. Identification and characterization of potential performance-related gut microbiotas in broiler chickens across various feeding trials. Appl. Environ. Microbiol. 2011, 77, 5868–5878. [Google Scholar] [CrossRef] [Green Version]
- Stanley, D.; Denman, S.E.; Hughes, R.J.; Geier, M.S.; Crowley, T.M.; Chen, H.; Haring, V.R.; Moore, R.J. Intestinal microbiota associated with differential feed conversion efficiency in chickens. Appl. Microbiol. Biotechnol. 2012, 96, 1361–1369. [Google Scholar] [CrossRef]
- Stanley, D.; Hughes, R.J.; Moore, R.J. Microbiota of the chicken gastrointestinal tract: Influence on health, productivity and disease. Appl. Microbiol. Biotechnol. 2014, 98, 4301–4310. [Google Scholar] [CrossRef]
- Pan, D.; Yu, Z. Intestinal microbiome of poultry and its interaction with host and diet. Gut Microbes 2014, 5, 108–119. [Google Scholar] [CrossRef]
- Kers, J.G.; Velkers, F.C.; Fischer, E.A.J.; Hermes, G.D.A.; Stegeman, J.A.; Smidt, H. Host and environmental factors affecting the intestinal microbiota in chickens. Front. Microbiol. 2018, 9, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stanley, D.; Geier, M.S.; Hughes, R.J.; Denman, S.E.; Moore, R.J. Highly variable microbiota development in the chicken gastrointestinal tract. PLoS ONE 2013, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deusch, S.; Tilocca, B.; Camarinha-Silva, A.; Seifert, J. News in livestock research—Use of Omics—Technologies to study the microbiota in the gastrointestinal tract of farm animals. Comput. Struct. Biotechnol. J. 2015, 13, 55–63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borda-Molina, D.; Seifert, J.; Camarinha-Silva, A. Current perspectives of the chicken gastrointestinal tract and its microbiome. Comput. Struct. Biotechnol. J. 2018, 16, 131–139. [Google Scholar] [CrossRef]
- Shang, Y.; Kumar, S.; Oakley, B.; Kim, W.K. Chicken gut microbiota: importance and detection technology. Front. Vet. Sci. 2018, 5. [Google Scholar] [CrossRef]
- Bäckhed, F.; Ley, R.E.; Sonnenburg, J.L.; Peterson, D.A.; Gordon, J.I. Host-bacterial mutualism in the human intestine. Science 2005, 307, 1915–1920. [Google Scholar] [CrossRef] [Green Version]
- Ley, R.E.; Turnbaugh, P.J.; Klein, S.; Gordon, J.I. Human gut microbes associated with obesity. Nature 2006, 444, 1022–1023. [Google Scholar] [CrossRef]
- Duncan, S.H.; Lobley, G.E.; Holtrop, G.; Ince, J.; Johnstone, A.M.; Louis, P.; Flint, H.J. Human colonic microbiota associated with diet, obesity and weight loss. Int. J. Obes. 2008, 32, 1720–1724. [Google Scholar] [CrossRef] [Green Version]
- Schwiertz, A.; Taras, D.; Schäfer, K.; Beijer, S.; Bos, N.A.; Donus, C.; Hardt, P.D. Microbiota and SCFA in lean and overweight healthy subjects. Obesity 2010, 18, 190–195. [Google Scholar] [CrossRef]
- Delzenne, N.M.; Cani, P.D. Interaction between obesity and the gut microbiota: Relevance in nutrition. Annu. Rev. Nutr. 2011, 31, 15–31. [Google Scholar] [CrossRef] [Green Version]
- Han, G.G.; Kim, E.B.; Lee, J.; Lee, J.Y.; Jin, G.; Park, J.; Huh, C.S.; Kwon, I.K.; Kil, D.Y.; Choi, Y.J.; et al. Relationship between the microbiota in different sections of the gastrointestinal tract, and the body weight of broiler chickens. Springerplus 2016, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- FAO. Probiotics in Animal Nutrition—Production, Impact and Regulation; FAO Animal Production and Health: Rome, Italy, 2016. [Google Scholar]
- Kageyama, A.; Benno, Y. Emendation of genus Collinsella and proposal of Collinsella stercoris sp. nov. and Collinsella intestinalis sp. nov. Int. J. Syst. Evol. Microbiol. 2000, 50, 1767–1774. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ricke, S. Perspectives on the use of organic acids and short chain fatty acids as antimicrobials. Poult. Sci. 2003, 82, 632–639. [Google Scholar] [CrossRef] [PubMed]
- Marteau, P.; Seksik, P.; Lepage, P.; Dore, J. Cellular and physiological effects of probiotics and prebiotics. Mini-Rev. Med. Chem. 2012, 4, 889–896. [Google Scholar] [CrossRef]
- Czepiel, J.; Dróżdż, M.; Pituch, H.; Kuijper, E.J.; Perucki, W.; Mielimonka, A.; Goldman, S.; Wultańska, D.; Garlicki, A.; Biesiada, G. Clostridium difficile infection: Review. Eur. J. Clin. Microbiol. Infect. Dis. 2019, 38, 1211–1221. [Google Scholar] [CrossRef] [Green Version]
- Moore, W.E.C.; Johnson, J.L.; Holdeman, L.V. Emendation of bacteroidaceae and butyrivibrio and descriptions of desulfomonas gen. nov. and ten new species in the genera desulfomonas, butyrivibrio, eubacterium, clostridium, and ruminococcus. Int. J. Syst. Bacteriol. 1976, 26, 238–252. [Google Scholar] [CrossRef] [Green Version]
- Ezaki, T.; Kawamura, Y.; Li, N.; Li, Z.Y.; Zhao, L.; Shu, S.E. Proposal of the genera Anaerococcus gen. nov., Peptoniphilus gen. nov. and Gallicola gen. nov for members of the genus Peptostreptococcus. Int. J. Syst. Evol. Microbiol. 2001, 51, 1521–1528. [Google Scholar] [CrossRef] [Green Version]
- Lawson, P.A.; Finegold, S.M. Reclassification of ruminococcus obeum as blautia obeum comb. nov. Int. J. Syst. Evol. Microbiol. 2015, 65, 789–793. [Google Scholar] [CrossRef]
- Ezaki, T.; Li, N.; Hashimoto, Y.; Miura, H.; Yamamoto, H. 16S ribosomal DNA sequences of anaerobic cocci and proposal of Ruminococcus hansenii comb. nov. and Ruminococcus productus comb. nov. Int. J. Syst. Bacteriol. 1994, 44, 130–136. [Google Scholar] [CrossRef]
- Liu, C.; Finegold, S.M.; Song, Y.; Lawson, P.A. Reclassification of Clostridium coccoides, Ruminococcus hansenii, Ruminococcus hydrogenotrophicus, Ruminococcus luti, Ruminococcus productus and Ruminococcus schinkii as Blautia coccoides gen. nov., comb. nov., Blautia hansenii comb. nov., Blautia hydroge. Int. J. Syst. Evol. Microbiol. 2008, 58, 1896–1902. [Google Scholar] [CrossRef]
- Rivière, A.; Selak, M.; Lantin, D.; Leroy, F.; De Vuyst, L. Bifidobacteria and butyrate-producing colon bacteria: Importance and strategies for their stimulation in the human gut. Front. Microbiol. 2016, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, H.; Wang, J.; He, T.; Becker, S.; Zhang, G.; Li, D.; Ma, X. Butyrate: A Double-Edged Sword for Health? Adv. Nutr. 2018, 9, 21–29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rios-Covian, D.; Salazar, N.; Gueimonde, M.; de los Reyes-Gavilan, C.G. Shaping the metabolism of intestinal bacteroides population through diet to improve human health. Front. Microbiol. 2017, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jacobson, A.; Lam, L.; Rajendram, M.; Tamburini, F.; Honeycutt, J.; Pham, T.; Van Treuren, W.; Pruss, K.; Stabler, S.R.; Lugo, K.; et al. A gut commensal-produced metabolite mediates colonization resistance to salmonella infection. Cell Host Microbe 2018, 24, 296–307. [Google Scholar] [CrossRef] [Green Version]
- El Hage, R.; Hernandez-Sanabria, E.; Arroyo, M.C.; Props, R.; Van De Wiele, T. Propionate-producing consortium restores antibiotic-induced dysbiosis in a dynamic in vitro model of the human intestinal microbial ecosystem. Front. Microbiol. 2019, 10. [Google Scholar] [CrossRef]
- Uzal, F.A.; Plattner, B.L.; Hostetter, J.M. Alimentary System. In Jubb, Kennedy and Palmer’s Pathology of Domestic Animals, 6th ed.; Elsevier: Amsterdam, The Netherlands, 2015; Volume 2, pp. 1–257.e2. [Google Scholar] [CrossRef]
- Rosenthal, M.E.; Rojtman, A.D.; Frank, E. Finegoldia magna (formerly Peptostreptococcus magnus): An overlooked etiology for toxic shock syndrome? Med. Hypotheses 2012, 79, 138–140. [Google Scholar] [CrossRef]
- Brook, I. Anaerobic Bacteria. In Infectious Diseases, 4th ed.; Cohen, J., Powderly, W.G., Opal, S.M., Eds.; Elsevier: Amsterdam, The Netherlands, 2017; pp. 1628–1644.e2. [Google Scholar] [CrossRef]
- Debrah, M.; Feingold, A.R. Anaerobic Cocci. In Principles and Practice of Pediatric Infectious Diseases, 4th ed.; Long, S.S., Ed.; Saunders: Philadelphia, PA, USA, 2012; pp. 988–990.e1. [Google Scholar] [CrossRef]
- Ley, R.E. Prevotella in the gut: Choose carefully. Nat. Rev. Gastroenterol. Hepatol. 2016, 13, 69. [Google Scholar] [CrossRef]
- Vartoukian, S.R.; Palmer, R.M.; Wade, W.G. The division “Synergistes. ” Anaerobe 2007, 13, 99–106. [Google Scholar] [CrossRef]
- Gardiner, B.J.; Tai, A.Y.; Kotsanas, D.; Francis, M.J.; Roberts, S.A.; Ballard, S.A.; Junckerstorff, R.K.; Kormana, T.M. Clinical and microbiological characteristics of eggerthella lenta bacteremia. J. Clin. Microbiol. 2015, 53, 626–635. [Google Scholar] [CrossRef] [Green Version]
- Shepherd, E.M.; Fairchild, B.D. Footpad dermatitis in poultry. Poult. Sci. 2010, 89, 2043–2051. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredients (g/100 g) | Starter (0–14 d) | Grower (15–28 d) | Finisher (29–42 d) |
|---|---|---|---|
| Corn | 42.17 | 34.96 | 12.73 |
| White corn | 0.00 | 0.00 | 15.00 |
| Wheat | 10.00 | 20.00 | 25.01 |
| Sorghum | 0.00 | 0.00 | 5.00 |
| Soybean meal | 23.11 | 20.63 | 17.60 |
| Expanded soybean | 10.00 | 10.00 | 13.00 |
| Sunflower | 3.00 | 3.00 | 3.00 |
| Corn gluten meal | 4.00 | 3.00 | 0.00 |
| Soybean oil | 3.08 | 4.43 | 5.48 |
| Dicalcium phosphate | 1.52 | 1.20 | 0.57 |
| Calcium carbonate | 0.91 | 0.65 | 0.52 |
| Sodium bicarbonate | 0.15 | 0.10 | 0.15 |
| Salt | 0.27 | 0.27 | 0.25 |
| Choline chloride | 0.10 | 0.10 | 0.10 |
| Lysine sulphate | 0.59 | 0.55 | 0.46 |
| Dl-methionine | 0.27 | 0.29 | 0.30 |
| Threonine | 0.15 | 0.14 | 0.14 |
| Xylanase | 0.08 | 0.08 | 0.08 |
| Phytase | 0.10 | 0.10 | 0.10 |
| Vitamin-mineral premix 1 | 0.50 | 0.50 | 0.50 |
| Proximate composition (g/100 g) | |||
| Dry matter | 88.57 | 88.65 | 88.64 |
| Protein | 22.70 | 21.49 | 19.74 |
| Lipid | 7.06 | 8.24 | 9.74 |
| Fiber | 3.08 | 3.04 | 3.07 |
| Ash | 5.85 | 5.17 | 4.49 |
| Lys | 1.38 | 1.29 | 1.21 |
| Ca | 0.91 | 0.80 | 0.59 |
| P | 0.63 | 0.57 | 0.46 |
| ME * (kcal/kg) | 3076 | 3168 | 3264 |
| Variables | A | B | C | D | p-Value |
|---|---|---|---|---|---|
| - n. | 9 | 10 | 9 | 10 | |
| Starter 0–14 d | |||||
| Chick body weight (g/bird) | 49.6 ± 0.7 | 50.1 ± 0.8 | 49.1 ± 0.9 | 49.2 ± 0.9 | 0.07 |
| Body weight (g/bird) | 470 ± 38.7 | 458.1 ± 30.2 | 472.8 ± 21.4 | 462.1 ± 28.2 | 0.70 |
| Daily weight gain (g/bird/d) 1 | 29.8 ± 2.8 | 29.1 ± 2.1 | 30.1 ± 1.3 | 29.5 ± 2.0 | 0.79 |
| Daily feed intake (g/bird/d) 1 | 39.5 ± 3.9 | 38.3 ± 2.5 | 40.4 ± 2.1 | 39.5 ± 2.5 | 0.44 |
| Feed intake (kg/bird) 1 | 0.55 ± 0.05 | 0.54 ± 0.04 | 0.57 ± 0.03 | 0.55 ± 0.03 | 0.44 |
| Feed conversion ratio 1 | 1.325 ± 0.05 | 1.318 ± 0.11 | 1.342 ± 0.05 | 1.340 ± 0.04 | 0.83 |
| Mortality (%) | 2.33 ± 2.22 | 0.83 ± 1.76 | 1.39 ± 2.95 | 0.00 | 0.10 |
| Grower 15–29 d | |||||
| Body weight (g/bird) | 1731 ± 61 | 1737 ± 63 | 1755 ± 46 | 1756 ± 35 | 0.65 |
| Daily weight gain (g/bird/d) 1 | 83.9 ± 3.0 | 84.7 ± 2.2 | 85.3 ± 2.3 | 85.8 ± 1.6 | 0.32 |
| Daily feed intake (g/bird/d) 1 | 131.7 ± 6.5 | 128.4 ± 12.5 | 131.2 ± 2.5 | 129.5 ± 2.9 | 0.76 |
| Feed intake (kg/bird) 1 | 1.98 ± 0.10 | 1.93 ± 0.19 | 1.97 ± 0.04 | 1.94 ± 0.04 | 0.76 |
| Cumulative feed intake (kg/bird) 1 | 2.53 ± 0.14 | 2.46 ± 0.20 | 2.53 ± 0.05 | 2.50 ± 0.06 | 0.61 |
| Feed conversion ratio 1 | 1.570 ± 0.07 | 1.519 ± 0.17 | 1.538 ± 0.04 | 1.509 ± 0.04 | 0.53 |
| Cumulative feed conversion ratio 1 | 1.506 ± 0.05 | 1.467 ± 0.14 | 1.487 ± 0.04 | 1.467 ± 0.03 | 0.67 |
| Mortality (%) | 0.00 | 1.67 ± 2.15 | 1.39 ± 2.08 | 0.83 ± 1.76 | 0.20 |
| Cumulative mortality (%) | 2.31 ± 2.20 | 3.33 ± 1.76 | 2.78 ± 4.17 | 0.83 ± 1.76 | 0.37 |
| Finisher 30–42 d | |||||
| Body weight (g/bird) | 3175 ± 98 | 3190 ± 93 | 3221 ± 95 | 3242 ± 83 | 0.40 |
| Daily weight gain (g/bird/d) 1 | 111.5 ± 4.6 | 110.7 ± 6.8 | 112.4 ± 4.7 | 114.2 ± 5.7 | 0.55 |
| Daily feed intake (g/bird/d) 1 | 201.5 ± 6.6 | 201.4 ± 5.5 | 199.8 ± 9.3 | 200 ± 5 | 0.92 |
| Feed intake (kg/bird) 1 | 2.62 ± 0.09 | 2.62 ± 0.07 | 2.60 ± 0.12 | 2.60 ± 0.07 | 0.92 |
| Feed conversion ratio 1 | 1.809 ± 0.07 | 1.823 ± 0.09 | 1.780 ± 0.09 | 1.753 ± 0.06 | 0.18 |
| Mortality (%) | 0.00 | 1.35 ± 2.16 | 0.51 ± 1.52 | 0.87 ± 1.83 | 0.34 |
| Overall experiment duration 0–42 d | |||||
| Chick body weight (g/bird) | 49.6 ± 0.7 | 50.1 ± 0.8 | 49.1 ± 0.9 | 49.2 ± 0.9 | 0.07 |
| Body weight (g/bird) | 3175 ± 98 | 3190 ± 93 | 3221 ± 95 | 3242 ± 83 | 0.40 |
| Daily weight gain (g/bird/d) 1 | 74.3 ± 2.3 | 74.6 ± 2.2 | 75.3 ± 2.2 | 75.9 ± 1.9 | 0.37 |
| Daily feed intake (g/bird/d) 1 | 120.7 ± 4.0 | 118.5 ± 4.7 | 119.3 ± 2.0 | 119.4 ± 2.2 | 0.61 |
| Feed intake (kg/bird) 1 | 5.15 ± 0.17 | 5.08 ± 0.21 | 5.13 ± 0.14 | 5.09 ± 0.09 | 0.79 |
| Feed conversion ratio 1 | 1.643 ± 0.05 | 1.625 ± 0.09 | 1.617 ± 0.04 | 1.596 ± 0.03 | 0.34 |
| Mortality (%) | 2.32 ± 2.2 | 4.58 ± 2.37 | 3.24 ± 4.55 | 1.67 ± 2.15 | 0.25 |
| Variables | A | D | p-value |
|---|---|---|---|
| n. | 9 | 10 | |
| Starter 0–14 d | |||
| Chick body weight (g/bird) | 49.6 ± 0.7 | 49.2 ± 0.9 | 0.57 |
| Body weight (g/bird) | 470 ± 38.7 | 462.1 ± 28.2 | 0.63 |
| Daily weight gain (g/bird/d) 1 | 29.8 ± 2.8 | 29.5 ± 2.0 | 0.77 |
| Daily feed intake (g/bird/d) 1 | 39.5 ± 3.9 | 39.5 ± 2.5 | 0.99 |
| Feed intake (kg/bird) 1 | 0.55 ± 0.05 | 0.55 ± 0.03 | 0.99 |
| Feed conversion ratio 1 | 1.325 ± 0.05 | 1.340 ± 0.04 | 0.44 |
| Mortality (%) | 2.33 ± 2.22 | 0.00 | < 0.01 |
| Grower 15–29 d | |||
| Body weight (g/bird) | 1731 ± 61 | 1756 ± 35 | 0.28 |
| Daily weight gain (g/bird/d) 1 | 83.9 ± 3.0 | 85.8 ± 1.6 | 0.09 |
| Daily feed intake (g/bird/d) 1 | 131.7 ± 6.5 | 129.5 ± 2.9 | 0.34 |
| Feed intake (kg/bird) 1 | 1.98 ± 0.10 | 1.94 ± 0.04 | 0.35 |
| Cumulative feed intake (kg/bird) 1 | 2.53 ± 0.14 | 2.50 ± 0.06 | 0.51 |
| Feed conversion ratio 1 | 1.570 ± 0.07 | 1.509 ± 0.04 | 0.03 |
| Cumulative feed conversion ratio 1 | 1.506 ± 0.05 | 1.467 ± 0.03 | 0.06 |
| Mortality (%) | 0.00 | 0.83 ± 1.76 | 0.17 |
| Cumulative mortality (%) | 2.31 ± 2.20 | 0.83 ± 1.76 | 0.29 |
| Finisher 30–42 d | |||
| Body weight (g/bird) | 3175 ± 98 | 3242 ± 83 | 0.13 |
| Daily weight gain (g/bird/d) 1 | 111.5 ± 4.6 | 114.2 ± 5.7 | 0.27 |
| Daily feed intake (g/bird/d) 1 | 201.5 ± 6.6 | 200 ± 5 | 0.57 |
| Feed intake (kg/bird) 1 | 2.62 ± 0.09 | 2.60 ± 0.07 | 0.57 |
| Feed conversion ratio 1 | 1.809 ± 0.07 | 1.753 ± 0.06 | 0.07 |
| Mortality (%) | 0.00 | 0.87 ± 1.83 | 0.17 |
| Overall experiment duration 0–42 d | |||
| Chick body weight (g/bird) | 49.6 ± 0.7 | 49.2 ± 0.9 | 0.37 |
| Body weight (g/bird) | 3175 ± 98 | 3242 ± 83 | 0.13 |
| Daily weight gain (g/bird/d) 1 | 74.3 ± 2.3 | 75.9 ± 1.9 | 0.11 |
| Daily feed intake (g/bird/d) 1 | 120.7 ± 4.0 | 119.4 ± 2.2 | 0.42 |
| Feed intake (kg/bird) 1 | 5.15 ± 0.17 | 5.09 ± 0.09 | 0.41 |
| Feed conversion ratio 1 | 1.643 ± 0.05 | 1.596 ± 0.03 | 0.01 |
| Mortality (%) | 2.32 ± 2.20 | 1.67 ± 2.15 | 0.82 |
| Variables | A | B | C | D |
|---|---|---|---|---|
| Number of birds | 213 | 207 | 206 | 224 |
| Score 0 (no lesion) (%) | 83.1 | 88.4 | 92.7 | 95.1 |
| Score 1 (moderate lesions) (%) | 10.8 | 4.8 | 4.4 | 4.0 |
| Score 2 (severe lesions) (%) | 6.1 | 6.8 | 2.9 | 0.9 |
| Chi-square test (p-value) | 0.07 | |||
| Variables | A | D |
|---|---|---|
| Number of birds | 213 | 224 |
| Score 0 (no lesion) (%) | 83.1 | 95.1 |
| Score 1 (moderate lesions) (%) | 10.8 | 4.0 |
| Score 2 (severe lesions) (%) | 6.1 | 0.9 |
| Chi-square test (p-value) | 0.02 | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Brugaletta, G.; De Cesare, A.; Zampiga, M.; Laghi, L.; Oliveri, C.; Zhu, C.; Manfreda, G.; Syed, B.; Valenzuela, L.; Sirri, F. Effects of Alternative Administration Programs of a Synbiotic Supplement on Broiler Performance, Foot Pad Dermatitis, Caecal Microbiota, and Blood Metabolites. Animals 2020, 10, 522. https://doi.org/10.3390/ani10030522
Brugaletta G, De Cesare A, Zampiga M, Laghi L, Oliveri C, Zhu C, Manfreda G, Syed B, Valenzuela L, Sirri F. Effects of Alternative Administration Programs of a Synbiotic Supplement on Broiler Performance, Foot Pad Dermatitis, Caecal Microbiota, and Blood Metabolites. Animals. 2020; 10(3):522. https://doi.org/10.3390/ani10030522
Chicago/Turabian StyleBrugaletta, Giorgio, Alessandra De Cesare, Marco Zampiga, Luca Laghi, Chiara Oliveri, Chenglin Zhu, Gerardo Manfreda, Basharat Syed, Luis Valenzuela, and Federico Sirri. 2020. "Effects of Alternative Administration Programs of a Synbiotic Supplement on Broiler Performance, Foot Pad Dermatitis, Caecal Microbiota, and Blood Metabolites" Animals 10, no. 3: 522. https://doi.org/10.3390/ani10030522